 $\operatorname {GL}(3)\times \operatorname {GL}(2)$ L-FUNCTIONS
$\operatorname {GL}(3)\times \operatorname {GL}(2)$ L-FUNCTIONS
In this paper, we prove uniform bounds for 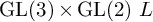 $\operatorname {GL}(3)\times \operatorname {GL}(2) \ L$-functions in the
$\operatorname {GL}(3)\times \operatorname {GL}(2) \ L$-functions in the 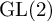 $\operatorname {GL}(2)$ spectral aspect and the t aspect by a delta method. More precisely, let
$\operatorname {GL}(2)$ spectral aspect and the t aspect by a delta method. More precisely, let 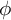 $\phi $ be a Hecke–Maass cusp form for
$\phi $ be a Hecke–Maass cusp form for 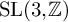 $\operatorname {SL}(3,\mathbb {Z})$ and f a Hecke–Maass cusp form for
$\operatorname {SL}(3,\mathbb {Z})$ and f a Hecke–Maass cusp form for 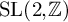 $\operatorname {SL}(2,\mathbb {Z})$ with the spectral parameter
$\operatorname {SL}(2,\mathbb {Z})$ with the spectral parameter 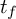 $t_f$. Then for
$t_f$. Then for 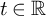 $t\in \mathbb {R}$ and any
$t\in \mathbb {R}$ and any 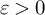 $\varepsilon>0$, we have Moreover, we get subconvexity bounds for
$\varepsilon>0$, we have Moreover, we get subconvexity bounds for 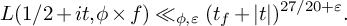 $$\begin{align*}L(1/2+it,\phi\times f) \ll_{\phi,\varepsilon} (t_f+|t|)^{27/20+\varepsilon}. \end{align*}$$
$$\begin{align*}L(1/2+it,\phi\times f) \ll_{\phi,\varepsilon} (t_f+|t|)^{27/20+\varepsilon}. \end{align*}$$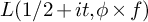 $L(1/2+it,\phi \times f)$ whenever
$L(1/2+it,\phi \times f)$ whenever 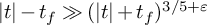 $|t|-t_f \gg (|t|+t_f)^{3/5+\varepsilon }$.
$|t|-t_f \gg (|t|+t_f)^{3/5+\varepsilon }$.

