
10 results
An improved method for isolating camallanid (Nematoda) spicules for scanning electron microscopy
-
- Journal:
- Journal of Helminthology / Volume 97 / 2023
- Published online by Cambridge University Press:
- 07 February 2023, e17
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Head-only electrical stunning and bleeding of African catfish (Clarias gariepinus): assessment of loss of consciousness
-
- Journal:
- Animal Welfare / Volume 13 / Issue 1 / February 2004
- Published online by Cambridge University Press:
- 11 January 2023, pp. 71-76
-
- Article
-
- You have access
- Export citation
Levels of arsenic, mercury and selenium in Clarias gariepinus from Sagua la Grande River, Cuba
-
- Journal:
- Annales de Limnologie - International Journal of Limnology / Volume 49 / Issue 2 / 2013
- Published online by Cambridge University Press:
- 13 June 2013, pp. 113-119
- Print publication:
- 2013
-
- Article
- Export citation
The quantitative requirement for α-tocopherol by juvenile African catfish, Clarias gariepinus Burchell
-
- Journal:
- Animal Science / Volume 65 / Issue 1 / August 1997
- Published online by Cambridge University Press:
- 02 September 2010, pp. 135-142
- Print publication:
- August 1997
-
- Article
- Export citation
Muscle and hepatic fatty acid profiles and α-tocopherol status in African catfish (Clarias gariepinus) given diets varying in oxidative state and vitamin E inclusion level
-
- Journal:
- Animal Science / Volume 64 / Issue 1 / February 1997
- Published online by Cambridge University Press:
- 02 September 2010, pp. 187-195
- Print publication:
- February 1997
-
- Article
- Export citation
Internal and external factors controlling reproduction in the African catfish, Clarias gariepinus
-
- Journal:
- Aquatic Living Resources / Volume 9 / Issue S1 / November 1996
- Published online by Cambridge University Press:
- 15 November 1996, pp. 45-58
- Print publication:
- November 1996
-
- Article
- Export citation
Effect of dietary protein on the nitrogen excretion and growth of the African catfish, Clarias gariepinus
-
- Journal:
- Aquatic Living Resources / Volume 8 / Issue 4 / October 1995
- Published online by Cambridge University Press:
- 15 October 1995, pp. 407-414
- Print publication:
- October 1995
-
- Article
- Export citation
-
The rates of growth and nitrogen efflux (total nitrogen and ammonia) of individual C. gariepinus ($\bar{x}~=~32.2$
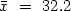 g; S.D. = 4.8 g) kept under 4 feeding regimens, following a 48 h imposed fast (phase l), were measured periodically. In phase 2 (35 d), groups A, B and C were fed a 49.75%, 45.55% or 41.10% protein diet respectively at a ration of 0.5% body weight (1 d−1) Group D were not fed. In phase 3 (25 d) all groups were fed the 41.10% diet. In phase 1, the ammonia efflux rates were lower than any of the values found in either phase 2 and 3. In phase 2, group A (49.75%) had a higher mean ammonia efflux rates than the other groups and ammonia comprised 60-100% of the total nitrogen efflux in all groups. Group D showed a direct relationship between ammonia efflux rate and length of fast. In phase 3, refeeding with the 41.1% protein diet caused the ammonia efflux rates of groups B and D to converge within 7 days to values no different from those of group C, but group A maintained a significantly higher mean value until day 25. During phase 2, the growth rates in B (45.55%) were greatest, but, none of the among groups differences were significant. Group D fish (unfed) lost approximately 30% of their initial weight during phase 2. Nitrogen efflux rates, notably ammonia, showed a pattern of excretion that was directly related to the protein content of the diet, but that the source of dietary protein, dietary energy and the total available energy also influenced nitrogen metabolism. The small differences in growth found were related to between diet differences in composition. The weight loss in the unfed group of fish was probably attributable to the utilisation of lipid and/or protein reserves as metabolic fuels.
g; S.D. = 4.8 g) kept under 4 feeding regimens, following a 48 h imposed fast (phase l), were measured periodically. In phase 2 (35 d), groups A, B and C were fed a 49.75%, 45.55% or 41.10% protein diet respectively at a ration of 0.5% body weight (1 d−1) Group D were not fed. In phase 3 (25 d) all groups were fed the 41.10% diet. In phase 1, the ammonia efflux rates were lower than any of the values found in either phase 2 and 3. In phase 2, group A (49.75%) had a higher mean ammonia efflux rates than the other groups and ammonia comprised 60-100% of the total nitrogen efflux in all groups. Group D showed a direct relationship between ammonia efflux rate and length of fast. In phase 3, refeeding with the 41.1% protein diet caused the ammonia efflux rates of groups B and D to converge within 7 days to values no different from those of group C, but group A maintained a significantly higher mean value until day 25. During phase 2, the growth rates in B (45.55%) were greatest, but, none of the among groups differences were significant. Group D fish (unfed) lost approximately 30% of their initial weight during phase 2. Nitrogen efflux rates, notably ammonia, showed a pattern of excretion that was directly related to the protein content of the diet, but that the source of dietary protein, dietary energy and the total available energy also influenced nitrogen metabolism. The small differences in growth found were related to between diet differences in composition. The weight loss in the unfed group of fish was probably attributable to the utilisation of lipid and/or protein reserves as metabolic fuels.
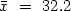
Genetic and nutritional influence on the total lipid fatty acid profile of Clarias gariepinus muscle
-
- Journal:
- Aquatic Living Resources / Volume 8 / Issue 4 / October 1995
- Published online by Cambridge University Press:
- 15 October 1995, pp. 415-421
- Print publication:
- October 1995
-
- Article
- Export citation
Controlled hatchery production of African catfish, Clarias gariepinus: the influence of temperature on early development
-
- Journal:
- Aquatic Living Resources / Volume 8 / Issue 4 / October 1995
- Published online by Cambridge University Press:
- 15 October 1995, pp. 431-438
- Print publication:
- October 1995
-
- Article
- Export citation
