



Introduction
Capsicum comprises a complex taxon (species and varieties) with a high genetic and phenotypic diversity. In this genus, 32 species have been described, but only five of them, Capsicum annuum, C. chinense, C. frutescens, C. baccatum, and C. pubescens, are domesticated (Baba et al., Reference Baba, Rocha, Gomes, de Fátima Ruas, Ruas, Rodríguez and Gonzalves2016). The Habanero pepper (Capsicum chinense Jacq.) is native to South America. However, it is cultivated worldwide, with Mexico standing out as one of the main producers. This species is particularly diversified in the Yucatan Peninsula, where local Habanero pepper fruits of many different shapes, sizes, and colours are found, due to the edaphoclimatic conditions of this region. These attributes placed it on the products list with the ‘Denomination of Origin’ of the Yucatan Peninsula and have contributed to the popularity of Habanero pepper in national and international markets (NOM, 2017), this is a traditional crop in Mexico, which is cultivated as a culinary product for export due to its flavour, typical aroma and strong itching, characteristics that have generated significant demand in the markets (Peña-Yam et al., Reference Peña-Yam, Muñoz-Ramírez, Avilés-Viñas, Canto-Flick, Pérez-Pastrana, Guzmán-Antonio, Santana-Buzzy, Aguilera-Cauich and Mijangos-Cortés2019b).
The Habanero fruit is an important ingredient in the cuisine of countries such as Mexico, China, Thailand and South Korea (Nass et al., Reference Nass, Souza, Ribeiro and Reifschneider2015), and is known for its high pungency (Ribeiro et al., Reference Ribeiro, Souza, Carvalho and Reifschneider2015) and unique aroma (Teodoro et al., Reference Teodoro, Alves, Ribeiro, Reis, Reifschneider, Fonseca and Agostini-Costa2013). It is also a rich source of vitamin C (Canto-Flick et al., Reference Canto-Flick, Balam-Uc, Bello-Bello, Lecona-Guzmán, Solís-Marroquín, Avilés-Viñas, Gómez-Uc, López-Puc, Santana-Buzzy and Iglesias-Andreu2008), capsaicinoids (De Jesús Ornelas-Paz et al., Reference De Jesús Ornelas-Paz, Martínez-Burrola, Ruiz-Cruz, Santana-Rodríguez, Ibarra-Junquera, Olivas and Pérez-Martínez2010; Sweat et al., Reference Sweat, Broatch, Borror, Hagan and Cahill2016; Jeeatid et al., Reference Jeeatid, Suriharn, Techawongstien, Chanthai, Bosland and Techawongstien2018), phenolic compounds (Menichini et al., Reference Menichini, Tundis, Bonesi, Loizzo, Conforti, Statti, De Cindio, Houghton and Menichini2009; Campos et al., Reference Campos, Gómez, Ordo and Ancona2013), carotenoids (Butcher et al., Reference Butcher, Crosby, Yoo, Patil, Ibrahim, Leskovar and Jifon2012; Giuffrida et al., Reference Giuffrida, Dugo, Torre, Bignardi, Cavazza, Corradini and Dugo2013; Gómez-García and Ochoa-Alejo, Reference Gómez-García and Ochoa-Alejo2013) flavonoids (Teodoro et al., Reference Teodoro, Alves, Ribeiro, Reis, Reifschneider, Fonseca and Agostini-Costa2013), and other secondary metabolites with antioxidant properties (Castro-Concha et al., Reference Castro-Concha, Tuyub-Che, Moo-Mukul, Vázquez-Flota and Miranda-Ham2014). These qualities make the Habanero pepper a key raw material for different industries (food, pharmaceutical, chemistry and cosmetology, among others), but the high demand for this product in the market has not yet been satisfied, mainly due to the absence of highly productive varieties and hybrids that preserve the attributes that consumers prefer.
Peppers (Capsicum spp.) are diploid and largely self-pollinated, so the hybridization method, as a source of new genetic combinations, has been widely used to improve most of the additive and nonadditive traits in this crop (Karim et al., Reference Karim, Rafii, Misran, Ismail, Khan and Chowdhury2021). Hybridization is a genetic improvement strategy that allows the transfer of genes of interest between species (interspecific) or within the same species (intraspecific) to develop genetically superior genotypes. Knowledge of the floral structure of a species and the nature of its reproductive process will facilitate the hybridization process (Zhang et al., Reference Zhang, Lin, Chen, Zhu and Chen2018). The Habanero pepper can selfpollinasy help, but the stigma and pollen are not receptive at the same time, and up to 90% outcrossing can occur. Studies related to anthesis, pollen viability and stigma receptivity, among others, are essential for the genetic improvement of this species (Hundal and Dhall, Reference Hundal and Dhall2004). Recent studies of flowering in Habanero pepper were addressed by Peña-Yam et al. (Reference Peña-Yam, Muñoz-Ramírez, Avilés-Viñas, Canto-Flick, Guzmán-Antonio and Santana-Buzzy2019a), who determined that before anthesis (flower bud), the stigma is already receptive, while the anthers are still closed in most of the genotypes studied; this characteristic ensures that self-fertilization cannot occur, despite the fact that the stigma is already receptive. This particularity in the floral biology of the Habanero pepper allows identifying this moment as the most appropriate to carry out manual pollination in this species in a hybridization programme.
In this way, it is possible to take advantage of combining ability and heterosis (hybrid vigour) during the selection of high-yielding genotypes with high fruit quality (Seneviratne and Kannangara, Reference Seneviratne and Kannangara2004; Pérez-Grajales et al., Reference Pérez-Grajales, González-Hernández, Peña-Lomelí and Sahagún-Castellanos2009). Heterosis is defined as the superiority of the F1 hybrid over its parents with respect to traits such as yield (1.99 Kg/Pta/1.31 Kg/Pta), growth rates, and other characteristics (Shull, Reference Shull1948). It has been widely used in agriculture to increase yield and broaden the adaptability of hybrid varieties (Singh, Reference Singh2016). Heterosis occurs when the hybrid is superior to the parental mean (average between two parents) of a specific variable. The term ‘heterosis’ describes the increased size and yield in crossbreds in comparison with their corresponding inbred; if there is no increase in these traits, there is no heterosis (Meyer et al., Reference Meyer, Torjek, Becher and Altmann2004). Heterobeltiosis expresses the betterment of the best parent.
Several breeding schemes have been used to evaluate variations, in general combining ability (GCA) and specific combining ability (SCA) of Capsicum (Amin et al., Reference Amin, Amiruzzaman, Ahmed and Ali2014). According to Sprague and Tatum (Reference Sprague and Tatum1942), the GCA estimates the average behaviour of a line in combinations with other lines and is related to genetic effects of additivity, and the SCA refers to the specific combinations with respect to the GCA of its parents, that is, it is the deviation of a cross with respect to the average behaviour of the parents and is related to dominance effects. The most used genetic designs in a breeding programme are biparental, multiple crosses, test crosses, line × tester (tester) and diallel designs. These designs allow determining the combining abilities of experiments of crosses and parental lines, in addition to understanding the inheritance of the investigated traits (Amin et al., Reference Amin, Amiruzzaman, Ahmed and Ali2014). One of the methods to assess SCA (line × tester) has been based on diallelic mating genetic designs, which basis were well described by Griffing (Reference Griffing1956). Most of the work done in the genus Capsicum to determine the magnitude of heterosis in hybrids has been done with C. annuum, and very few reports exist for other pepper species. Recently Peña-Yam (Reference Peña-Yam2020) and Ramírez (Reference Ramírez2020) reported obtaining both high yield and quality fruits F1 hybrids of Habanero pepper, under greenhouse conditions, for which they used 10 genotypes belonging to the germplasm preserved in Laboratory 9 of the Plant Biochemistry and Molecular Biology Unit (UBBMP), at Yucatan Scientific Research Center (CICY).
There is a growing and unsatisfied demand for Habanero pepper in the national and international market; however, there are few improvement programs and a high risk of genetic erosion of the varieties. For this reason, the researchers at Laboratory 9, have worked to generate improved varieties and more productive F1 hybrids. To do this, they rely on the estimation of statistical genetic parameters, which provide information on the genetic variation existing in the population, and thus support the progress of the selection process. The production of hybrids is an alternative that offers the producer a high yield potential, good climatic adaptation and resistance to the main diseases, as well as better fruit quality because it is superior to the characteristics of its parents. Therefore, a sustainable and competitive varietal creation is necessary to achieve new hybrids with different commercial purposes (Peña-Yam et al., Reference Peña-Yam, Muñoz-Ramírez, Avilés-Viñas, Canto-Flick, Guzmán-Antonio and Santana-Buzzy2019a).
With this background, the objective of this study was to identify parents with high yield potential to obtain new commercial F1 hybrids for fresh consumption.
Materials and methods
Ten genotypes of Habanero pepper (C. chinense Jacq) were selected from the germplasm conserved in UBBMP at CICY, as parents of F1 hybrids. The selection of genotypes for this study was based on their outstanding fruit agronomic characteristics, on evaluations carried out in previous studies, such as yield between 1.48–4.75 Kg/Pta, number of fruits per plant between 109–522 fruits, and average weight from 6.75 to 16.68 g. The selected genotypes were ASB-02, NEK-03, RNJ-04, RHC-05, NBA-06, AKN-07, RES-08, MSB-09, RKI-10 and NKA-11. The management conditions given to the genotypes are described below and were the same followed in the evaluation of the hybrids. The investigation was conducted in a greenhouse, at 20–35°C, with 55–75% of relative humidity and cycles of natural light (approximately, 11 h of light, 13 h of darkness), located in facilities at CICY, at 20° 58′2.53” North Latitude and 89° 35′33.30” West Longitude, and at an altitude of 10 masl (SMN, 2020).
Evaluation of the hybrids (F1)
Seeds from parents and hybrids were disinfected with commercial sodium hypochlorite (Cloralex 99.9%) for 10 min and sown in 200 well polystyrene trays to germinate. As a substrate, Peat Moss Sunshine (mix 3) (Sol®) was used. After sowing, trays were covered with black plastic, until the seeds started to germinate. The transplant was carried out in black bags (40 × 40 cm) containing a substrate composed of a 3:1 mix of red soil and Sunshine. To promote the development of the root system, Hakaphos® 13-40-13 commercial fertilizer (1 g/l) was applied once a week, during the development of plantlets. Triple 18 fertilizer (3 g/l) was applied weekly to enhance vegetative growth. Agronomic management of the plants followed the guidelines for greenhouse Habanero pepper cultivation (Tun, Reference Tun2001; Villa et al., Reference Villa Castorena, Catalán Valencia, Inzunza Ibarra, Román López, Macias Rodríguez and Cabrera Rodarte2014). The distance between plants and between lines was 40 and 80 cm, respectively.
Genetic mating design
The crosses were conducted manually from October 2018 to April 2019. From the germplasm, 26 genotypes were used, with 10 parents: two male testers (RNJ-04, RES-08), and 8 female lines (ASB-02, NEK-03, RHC-05, NBA-06, AKN-07, MSB-09, RKI-10, NKA-11). Sixteen hybrids were obtained: ASB-02 × RNJ-04, NEK-03 × RNJ-04, RHC-05 × RNJ-04, NBA-06 × RNJ-04, AKN-07 × RNJ-04, MSB-09 × RNJ-04, RKI-10 × RNJ-04, NKA-11 × RNJ-04, ASB-02 × RES-08, NEK-03 × RES-08, RHC-05 × RES-08, NBA-06 × RES-08, AKN-07 × RES-08, MSB-09 × RES-08, RKI-10 × RES-08, NKA-11 × RES-08. The fruits of the hybrid progeny (F1) of each cross were harvested when the fruit was completely ripe, in April 2021. Seeds were extracted from each fruit separately and placed in small Petri dishes (65.7 × 14.7 mm high), which were then placed in a 36°C oven for 72 h. After this time, seeds from each cross were stored in kraft paper envelopes (3.0 × 1.5 cm) and kept at 24–25°C until sowing for evaluation.
The assessed variables were: fruit length (FL): Measured at the longest point; an average of 10 ripe fruits in the second harvest, fruit diameter (FD): Measured at the widest point, and an average of 10 ripe fruits in the second harvest; thickness of the pericarp (TP): Average thickness of 10 ripe fruits of the second harvest; the number of fruits per plant (NFP): Average number of fruits in each harvest per plant; average fruit weight (AFW): Average weight of 10 ripe fruits of the second harvest; and yield per plant (YP): Average fruit yield on 10 plants, as described by the IPGRI Capsicum Genus Descriptors Manual (IPGRI, 1995).
Statistical analysis
Eight genotypes were used as female parents (ASB-02, NEK-03, RHC-05, NBA-06, AKN-07, MSB-09, RKI-10, NKA-11), and two genotypes were used as testers (RNJ-04 and RES-08). The ten parents were crossed according to the Line × Tester mating design reported by Kempthorne (Reference Kempthorne1957), the Griffing's method 2 model I (fixed effect) (Griffing, Reference Griffing1956). Genetic analysis was performed under the model proposed by Kempthorne and Curnow (Reference Kempthorne and Curnow1961), and described by Singh and Chaudhary (Reference Singh and Chaudhary1985).
The experiment was performed in a random block design with four repetitions. A total of six plants, of each genotype, were evaluated from each of the harvests carried out. Based on the design, and analysis of variance was conducted, and the means were compared using Tukey's HSD multiple range test (P ≤ 0.05) with the statistical package Statgraphics plus 5.0 (Corp, Reference Corp2000).
General and specific combining ability, heterosis, and heterobeltiosis
The GCA and SCA were determined following the methodology of Jenkins and Brunson (Reference Jenkins and Brunson1932). Standard errors of the effects of GCA and SCA were estimated as described by Singh (Reference Singh1973). The decomposition of the effect of genotypes in parents (P), crosses (C) and the interaction P vs C, was done as indicated by Hallauer and Miranda (Reference Hallauer and Miranda1988).
According to Martínez-Martínez et al. (Reference Martínez-Martínez, Méndez-Infante, Castañeda-Aldaz, Vera-Guzmán, Chávez-Servia and Carrillo-Rodríguez2014), heterosis (H) was estimated using the mean of the parents and expressed on a percentage scale. The significance of heterosis (H) was determined by Tukey's test with the statistical package Statgraphics plus 5.0. To identify hybrids with superior performance, heterobeltiosis (Hb) (if the vigour of the hybrid exceeds the expression of the best of its parents) was determined according to Robles (Reference Robles1986).
Results
All the agronomic variables recorded showed highly significant differences at P ≤ 0.05 (Table 1). The effect of genotypes, the effect of the crosses, and the effect of parents was significant in all the variables analysed, highlighting the AFW, NFP, and YP. The interaction P vs C also had a behaviour similar to the previous ones, which implies the existence of heterosis. The mean squares for the GCA were of greater magnitude than those of SCA in the variables studied, which suggests that the effects of the GCA contribute more to the genetic variation of the yield components in the genotypes than the effects of SCA. The GCA differed significantly (P ≤ 0.05) in fruit yield, the number of fruits per plant, and average fruit weight. For SCA, significant effects (P ≤ 0.05) were found in the number of fruits per plant, and the average weight of the fruit, which indicates the presence of nonadditive genetic action (dominance) in these characters. As a result, the range of variation was obtained in the CV values (2.30–11.71%) for each of the characteristics evaluated. The highest values corresponded to NFP (10.34%) and YP (11.71%), while the lowest values were for FL (3.68%), FD (2.30%), TP (4.11%) and AFW (3.31%).
Table 1. Analysis of the sources of variation and mean squares of the Line × Tester design for the Habanero pepper variables assessed
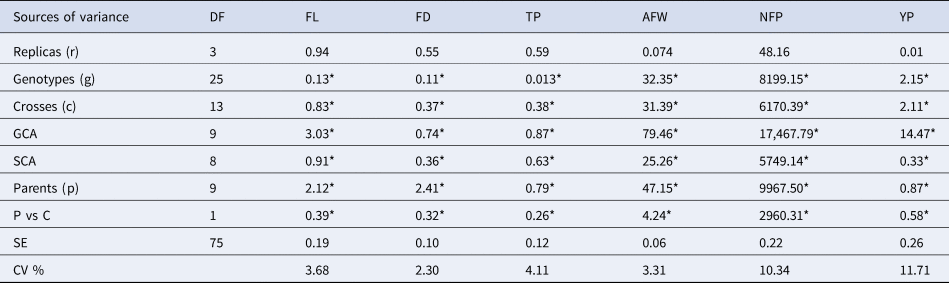
*Tukey's P ≤ 0.05; DF, Degrees of freedom; FL, Fruit Length (cm); FD, Fruit Diameter (cm); TP, thickness of the pericarp (mm); NFP, Number of fruits per plant; AFW, Average Fruit Weight (g); and YP, Yield per plant (Kg/Pta); GCA refers to parents; SCA refers to crosses; SE, Error Standard; CV, Coefficient of variation in percentage (%).
There were highly significant differences between genotypes in all the variables analysed (Table 2). The hybrid vigour, which is the expression of heterosis, can be seen in the average values of the hybrids, which exceeded the average values of their parents and the average of the experiment, for the variables FL, FD, NFP and YP, the opposite with what was observed for the FT and AFW variables. Regarding FL, the RNJ-04 parent used as a tester presented the highest value (5.70 cm) with respect to the rest of the parents, while the ASB-02 × RNJ-04 hybrid with 6.38 cm and the MSB-09 × RNJ-04 hybrid with 6.53 cm, surpassed the rest, for the same variable. For the FD variable, the parent ASB-02 reached the highest value with 4.05 cm; while the MSB-09 × RES-08 and ASB-02 × RNJ-04 hybrids presented the highest values (4.28 and 4.23 cm, respectively). In the TP variable, the parents ASB-02 (3.50 mm), NBA-06 (3.38 mm), and NKA-11 (3.50 mm) reached the highest values, as well as the ASB-02 × RNJ-04 (3.30 mm) and NEK-03 × RES-08 (3.18 mm). For the AFW variable, six parents and three hybrids presented values greater than 11 g, differing significantly from the rest of the genotypes. Parent RNJ-04 had the lowest mean fruit weight at 6.25 g. Regarding the NFP variable, the parents NEK-03 (199.25), RKI-10 (207.75) and NKA (193.50) showed the best results, while the RKI-10 × RNJ-04 hybrid presented the highest with 481.75 fruits by plants, differing significantly from the rest of the hybrids studied. Regarding the parents used as testers, RNJ-04 stood out for FL and NFP, while RES-08 for AFW.
Table 2. Comparison of the mean values of the six fruit variables studied in the ten parents and the 16 hybrids of Habanero pepper obtained
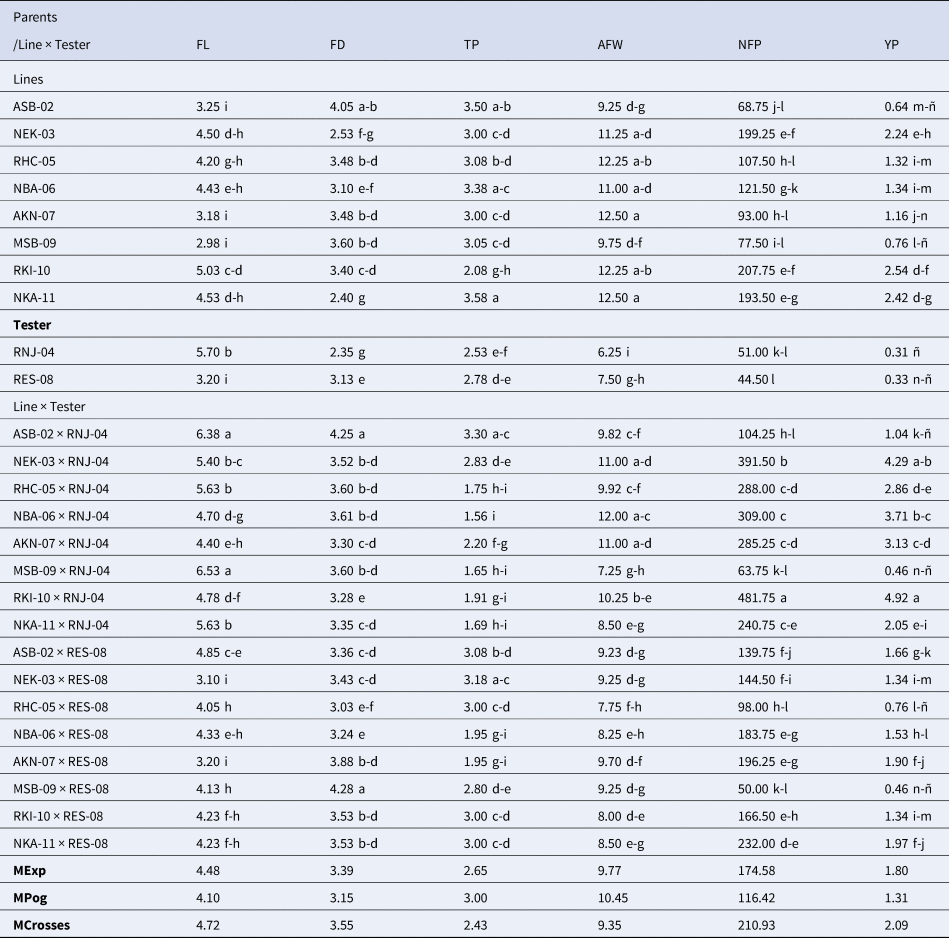
*Means with the same letter have no significant differences (Tukey's Studentized range test, P ≤ 0.05); MExp denotes the mean of the experiment; MPog denotes the mean of the parents and MCrosses denotes the mean of the crosses.
Regarding the yield variable, the parents with the best results were: NEK-03, RKI-10 and NKA-11 with values of 2.24, 2.54 and 2.42 Kg/Pta, respectively, while the RKI-10 × RNJ-04 hybrid produced 4.92 Kg/Pta and NEK-03 × RNJ-04 yielded 4.29 Kg/Pta, which allowed predicting yields of 98.4 and 85.8 tons/ha, respectively. The RKI-10 × RNJ-04 hybrid presented the highest value in the NFP variable, determining its high performance, while the AFW variable was the component with the greatest impact on the performance of the NEK-03 × RNJ-04 hybrid. Therefore, NFP and AFW were the variables that most influenced performance. The genetic difference between both parents could have promoted the high productive potential in these hybrids. There were also other promising hybrids, that showed good behaviour in yield, with values higher than 3.00 Kg/Pta, differing from the rest of the hybrids evaluated, these were: NBA-06xRNJ-04 with 3.71 Kg/Pta, which had the highest average weight of fruits (12.0 g), the largest number of fruits per plant (309.0) and large fruit size (FL 4.70 cm and FD 3.61 cm) and AKN-07xRNJ-04 hybrid with 3.13 Kg/Pta, 285.25 fruits per plant and average fruit weight of 11 g.
General and specific combining ability, heterosis, and heterobeltiosis
The effects of the estimated values of the GCA for the 10 Habanero pepper parents (Table 3) showed that most of the parents had high and positive values in all the analysed variables. Estimations of the parents’ GCA showed the best values for each fruit variable were: RNJ-04 for FL (1.60), ASB-02 for FD (0.87), NKA-11 for TP (0.56) and YP (2.18), RKI-10 for NFP (82.83) and RES-08 for AFW (2.05). The RES-08 also showed a high value in NFP (71.93). Furthermore, some of the parents showed notable effects of GCA in more than one variable such as ASB-02 for FD (0.87) and TP (0.47); NEK-03 for NFP (82.83) and YP (1.0); RES-08 for NFP (71.93) and AFW (2.95); RKI-10 for NFP (91.33) and YP (2.04); and finally NKA-11, with the highest values for TP (0.56), NFP (77.08), AFW (2.05) and YP (2.18) respectively, the latter being the parent with the best GCA.
Table 3. General combining capacity (ACG) in the six fruit variables for the 10 progenitors of Habanero pepper used in the study
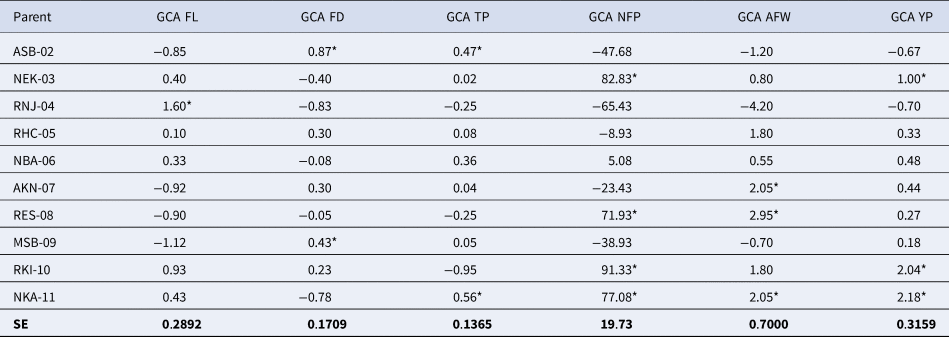
*Tukey's P ≤ 0.05 significant; GCA refers to parents; SE, standard error of the difference of parents means; FL, Fruit length (cm); FD, Fruit Diameter (cm); TP, thickness of the pericarp (mm); NFP, Number of fruits per plant; AFW, Average Fruit Weight (g), and YP, Yield per plant (Kg/Pta).
The SCA effects’ estimates in the 16 hybrids (Table 4) showed high and positive values. The best hybrids for each fruit variable were: ASB-02xRNJ-04 and RKI-10xRNJ-04 with values of 1.04 and 1.02 for FL respectively; NKA-11xRNJ-04 and RKI-10xRES-08 for FD, both with a value of 1.45; ASB-02xRNJ-04 for TP (0.54); ASB-02xRNJ-04 and NBA-06xRNJ-04 with the best AFW values (6.19 and 6.69), respectively. Other values to consider in this same variable are in the NEK-03xRNJ-04, RHC-05xRNJ-04, AKN-7xRNJ-04, and RKI-10xRNJ-04 hybrids with values of 5.44, 3.19, 4.19 and 2.56, respectively. For NFP, five hybrids showed the best results with values between 174.66 and 326.41, whereas the RKI-10xRNJ-04 hybrid showed the highest value for this variable, with 326.41 fruits, and the NBA-06xRNJ-04 hybrid for YP (2.90). The NBA-06 × RNJ-04 hybrid obtained the highest SCA values for the YP variable and its components, although other hybrids such as NEK-03xRNJ-04 (0.12) and RKI-10xRNJ-04 (0.83) also had outstanding values for the YP variable and that could be considered in similar selection programs.
Table 4. Specific combining capacity (SCA) and heterosis (H) values in 16 Habanero chili hybrids for the six fruit variables studied
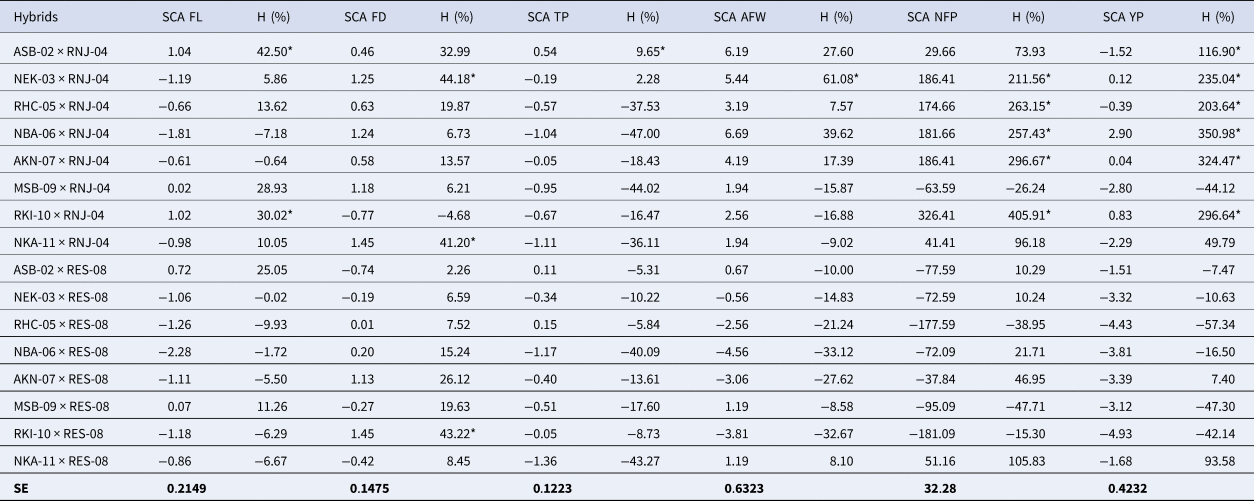
*Tukey's P ≤ 0.05 significant; SCA to crosses; H, Heterosis (%); SE, Standard error of SCA effects.
Regarding the results of heterosis (H) (Table 4), all the variables analysed showed high and positive values. The highest percentages of heterosis were recorded for NFP and YP. These variables could be improved more efficiently by breeding methods that take advantage of the effects of non-additive gene action, such as hybridization. For the FL variable, the best values were reached by the ASB-02xRNJ-04 (42.50%) and the RKI-10xRNJ-04 (30.02%) hybrids; for FD, there were three (NEK-03 × RNJ-04, NKA-11 × RNJ-04, RKI-10xRES-08) hybrids that obtained the best values with 44.18, 41.20 and 43.22%, respectively. The ASB-02xRNJ-04 hybrid obtained 9.65% for the TP variable. The NEK-03 × RNJ-04 hybrid was the best value for AFW (61.08%). Five hybrids achieved the best values for both NFP (174.66–405.91%) and YP (203.64–350.98%). The RKI-10 × RNJ-04 hybrid obtained the highest percentage of heterosis for NFP (405.91%) and the NBA-06 × RNJ-04 hybrid for YP (350.98%).
The percentages of heterobeltiosis (Hb) obtained among the Habanero pepper hybrids are shown in Table 5. Hb percentages greater than 100% were found for NFP and YP. For both the NFP and YP fruit variables, five hybrids obtained the best values in a range between 153.71–346.90% for NFP and between 119.69–274.34% for YP. In the AFW variable, three hybrids showed values between 9.03–10.61%. The RKI-10 × RNJ-04 hybrid stands out with the highest values for NFP (346.90%) and for YP (274.345%) and the ASB-02 × RNJ-04 hybrid for AFW (10.61%). The component that contributed the most to performance was NFP. According to the results of this study, we can highlight that the hybrid obtained using masculine tester RNJ-04 had a significant superiority over the rest of the crosses, in terms of SCA, H, and Hb, for most of the variables analysed, as well as for YP and its components. The NEK-03 × RNJ-04, NBA-06 × RNJ-04, RKI-10 × RNJ-04, and AKN-07 × RNJ-04 F1 hybrids were selected and recommended for inclusion in a programme of hybrid seed production.
Table 5. Percentages of heterobeltiosis (Hb) obtained by the Habanero pepper hybrids for the yield variables and their components
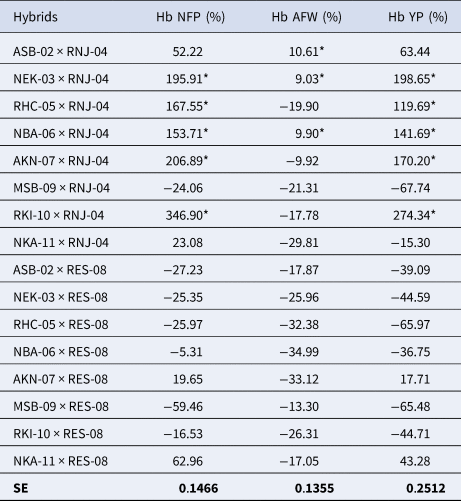
*Tukey's P ≤ 0.05 significant; Hb, heterobeltiosis (%); NFP, number of fruits per plant; AFW, Average fruit weight (g), and YP, Yield per plant (Kg/Pta).
Discussion
There were highly significant differences for each agronomic variable in this work, which demonstrates the wide variability of the study material. The effects of both genotype cross, and parents were high for yield, weight, and the number of fruits per plant. Similar results were reported by Gomide et al. (Reference Gomide, Maluf and Gomes2003) for Capsicum annuum fruit yield and Pech-May et al. (Reference Pech-May, Castañón-Nájera, Tun-Suárez, Mendoza-Elos, Mijangos-Cortés, Pérez-Gutiérrez and Latournerie-Moreno2010) on Sweet pepper for yield and its components. That is, the evaluated characters of the germplasm were associated more with additive effects than with non-addictive ones, as has been reported in ¨Serrano¨ by Martínez-Martínez et al. (Reference Martínez-Martínez, Méndez-Infante, Castañeda-Aldaz, Vera-Guzmán, Chávez-Servia and Carrillo-Rodríguez2014) and in chili (C. annuum) by Nandadevi and Hosamani (Reference Nandadevi and Hosamani2003). GCA and SCA differed significantly (P ≤ 0.05) in yield and its components, indicating the presence of nonadditive genetic action (dominance) in these traits. These results coincide with those reported in Sweet pepper by Pech-May et al. (Reference Pech-May, Castañón-Nájera, Tun-Suárez, Mendoza-Elos, Mijangos-Cortés, Pérez-Gutiérrez and Latournerie-Moreno2010) but they differ in the weight of the fruit from that reported in ‘Serrano’ chili by Martínez et al. (Reference Martínez, Dorantes, Ramírez, de la Rosa and Pozo2005), who found the dominance effects were what determined this variable response; and it coincides with those of Lippert (Reference Lippert1975), who found that additive effects were more important than dominance for fruit weight.
According to Martínez et al. (Reference Martínez, Dorantes, Ramírez, de la Rosa and Pozo2005), the estimation of the CV provides the degree of variation in relation to the average of a given characteristic and shows the variability present within it, as well as its possibilities for improvement. This allows us to infer that the characteristics evaluated during our work were adequate to obtain substantial advances in the selection processes. Similar recommendations have been described by Meshram and Mukewar (Reference Meshram and Mukewar2011) and Turgut (Reference Turgut2003). Our results did not show a wide range in the CV values (2.30–11.71%) for the variables evaluated, but they helped to find wide morphological variability in the genotypes tested.
Peña-Yam et al. (Reference Peña-Yam, Muñoz-Ramírez, Avilés-Viñas, Canto-Flick, Pérez-Pastrana, Guzmán-Antonio, Santana-Buzzy, Aguilera-Cauich and Mijangos-Cortés2019b) and Amin et al. (Reference Amin, Amiruzzaman, Ahmed and Ali2014) observed highly significant differences among the genotypes under their studies, indicating that there was a wide range of variability among them. According to Leal and Porras (Reference Leal and Porras1998), the differences between hybrids result from the variability present among the parents under study, which allows the production of hybrids with different phenotypes. In this sense, Luna-Ortega et al. (Reference Luna-Ortega, García-Hernández, Valdez-Cepeda, Gallegos Robles, Preciado-Rangel, Guerrero-Guerrero and Espinoza-Banda2013) have indicated that significant differences (P ≤ 0.01) in the male × female interaction, can be attributed to the high genetic variability present between the parents, as well as to the effects of different combinations between the parents involved in crosses. Luna-García et al. (Reference Luna-García, Robledo-Torres, Vásquez-Badillo, Ramírez-Godina and Mendoza-Villarreal2018) studied hybridization between several types of peppers and stated there is enough genetic variability to develop superior varieties, given the significant differences between hybrids and parents, in terms of yield and its components. Hybrid vigour, which is the expression of heterosis, can be seen in the mean values of the hybrid, that exceeded the mean values of their parents, and the mean of the experiment for the variables FL, FD, NFP and YP. This contrasts with there was observed for the variables TP and AFW. In general, these results corroborate what has been reported by different authors, on the superiority of F1 obtained by crossing the less related parents, instead of the average parents of the population (Seneviratne and kannangara, Reference Seneviratne and Kannangara2004) and that hybrid vigour exploitation will have allowed the expression of high yield potential or other traits of economic interest (Falconer and Mackay, Reference Falconer and Mackay1996).
According to SIAP Servicio de Información Agroalimentaria y Pesquera de México (2009) report, under greenhouse conditions, the density of Habanero pepper plants is 2.4/m2, producing between 7 and 12 Kg/m2 of commercial fruit, which indicates that each plant yields, on average, 3 to 4.5 Kg/Pta of Habanero peppers. One would expect around 190 tons/ha. SIAP reported yields of 85.5 tons/ha (Peña-Yam et al., Reference Peña-Yam, Muñoz-Ramírez, Avilés-Viñas, Canto-Flick, Pérez-Pastrana, Guzmán-Antonio, Santana-Buzzy, Aguilera-Cauich and Mijangos-Cortés2019b) when evaluating 13 varieties of peppers (C. annuum), and found that ‘Orion hybrid’ showed the highest average fruit weight (178.7 g), good size (9.2 cm diameter and 7 cm length), best yield per plant (1.9 Kg/Pta) and per plot (11.5 Kg/m2). On the other hand, Moreno Pérez et al. (Reference Moreno Pérez, Mora Aguilar, Sánchez Castillo and García Pérez2011) found that the ‘Magno hybrid’ had the lowest yield (5.7 Kg/m2). In this work, two hybrids were obtained with a yield higher than 85.8 tons/ha. The hybrids obtained showed a significant increase in yield and in the number of fruits per plant, as well as the average weight of the fruit. These results corroborated those reported by Pech-May et al. (Reference Pech-May, Castañón-Nájera, Tun-Suárez, Mendoza-Elos, Mijangos-Cortés, Pérez-Gutiérrez and Latournerie-Moreno2010) who recommend the use of hybridization as a breeding method in C. annuum to increase the yield and the number of fruits per plant but not to improve characters, such as the length and width of the fruit, the thickness of the pericarp, or the average weight of the fruit.
In this study, six of the ten parents used, reached high and positive values for GCA in all fruit variables analysed. Parent NKA-11 obtained the highest values, which allows it to be recommended for future improvement works in this crop. In this sense, different authors such as Peña-Yam et al. (Reference Peña-Yam, Muñoz-Ramírez, Avilés-Viñas, Canto-Flick, Pérez-Pastrana, Guzmán-Antonio, Santana-Buzzy, Aguilera-Cauich and Mijangos-Cortés2019b) and Ramírez (Reference Ramírez2020) obtained similar estimated GCA values for the Habanero pepper parental lines. The most outstanding lines were ASBC-09 for FW, PT, and AF; RNJ-04 for FL and NFP, and RKI-01 for RP. Pech-May et al. (Reference Pech-May, Castañón-Nájera, Tun-Suárez, Mendoza-Elos, Mijangos-Cortés, Pérez-Gutiérrez and Latournerie-Moreno2010) and Zewdie et al. (Reference Zewdie, Bosland and Steiner2001) observed positive GCA values are the expression of variability present in parents, that can be transmitted to their offspring. Since GCA is associated with additive gene action in the inheritance of traits, hybridization can contribute to the improvement of a particular trait (Martínez et al., Reference Martínez, Dorantes, Ramírez, de la Rosa and Pozo2005). This allows the selection of plants that combine superior traits, to those of their parents, and contributes to predicting crosses with higher productive potential. Pech-May et al. (Reference Pech-May, Castañón-Nájera, Tun-Suárez, Mendoza-Elos, Mijangos-Cortés, Pérez-Gutiérrez and Latournerie-Moreno2010) reported higher GCA values for total fruit yield in P2, P3 and P4 parents, which showed the highest positive values (20.9, 41.5 and 19.6 Kg/Pta, respectively). On the other hand, Khalil and Hatem (Reference Khalil and Hatem2014) working with peppers (C. annuum) indicated that, based on estimates of GCA values, they were able to identify the best parental lines for each trait. The selection of parents with good overall combining ability is a primary requirement for a breeding programme's success, especially to increase heterosis.
There were found high and positive SCA values in all variables for the studied hybrids. Five of these hybrids presented the highest SCA value estimates. The best hybrid with high SCA estimates for yield variable and its components was NBA-06 × RNJ-04. Also, other lines were of interest for their SCA values for the yield variable and could be considered in future selection programs. Similar results were reported by Peña-Yam et al. (Reference Peña-Yam, Muñoz-Ramírez, Avilés-Viñas, Canto-Flick, Pérez-Pastrana, Guzmán-Antonio, Santana-Buzzy, Aguilera-Cauich and Mijangos-Cortés2019b) in Habanero pepper for the SCA of the yield variable, with values between 0.24 and 0.50 Kg/Pta. In addition, Pech-May et al. (Reference Pech-May, Castañón-Nájera, Tun-Suárez, Mendoza-Elos, Mijangos-Cortés, Pérez-Gutiérrez and Latournerie-Moreno2010) found significant effects of SCA for yield in different crosses carried out in sweet pepper (C. annuum) and obtained values of 0.77–0.33 Kg/Pta. Peña-Lomelí et al. (Reference Peña-Lomelí, Molina-Galán, Cervantes-Santana, Márquez-Sánchez, Sahagún-Castellanos and Ortiz-Cereceres1998) evaluated intervarietal heterosis in Mexican husk tomatoes (Physalis ixocarpa) and found significant GCA effects for the three variables evaluated (NFP, AFW, and YP), as well as significant SCA effects for the AFW variable. Based on that, they suggested additive effects were more important than nonadditive effects for the variables NFP and YP. Khalil and Hatem (Reference Khalil and Hatem2014) working with peppers, reported estimated high SCA values similar to those in our work. Ahmed et al. (Reference Ahmed, Khan and Tank1997) suggested that the most appropriate breeding methods, to take advantage of favourable alleles, can be defined using parents and their crosses with high GCA and SCA values.
From a practical perspective, hybrid vigour is more important when the F1 hybrid is superior to the best parent. Nowadays, heterosis is widely used in the agricultural production of different crops, such as rice, maize, vegetables, and some perennial plants (Yu et al., Reference Yu, Gu, Zhang, Dong, Miao, Gebretsadik and Bo2021; Zhang et al., Reference Zhang, Huang and Han2021) According to Wu et al. (Reference Wu, Liu, Zhang and Gu2021), heterosis is an indicator of the degree of genetic relationship between the parents, and its use has gradually improved the yield, quality and resistance to disease of animals and plants, thus increasing social and economic benefits of agriculture. In our study, five hybrids presented heterosis values above 100% for NFP, and six hybrids for YP. According to Cabrera (Reference Cabrera2016), if these heterosis values are above 100%, it can be stated that the two parents are genetically different. High values of yield heterosis have been detected in Capsicum. Ahmed and Muzafar (Reference Ahmed and Muzafar2000) reported an average heterosis value of 174.52%, Seneviratne and Kannangara (Reference Seneviratne and Kannangara2004) of up to 154%, and Meshram and Mukewar (Reference Meshram and Mukewar2011) of 157%. Similar results were recorded by Peña-Yam et al. (Reference Peña-Yam, Muñoz-Ramírez, Avilés-Viñas, Canto-Flick, Pérez-Pastrana, Guzmán-Antonio, Santana-Buzzy, Aguilera-Cauich and Mijangos-Cortés2019b) in Habanero pepper: 504.14% heterosis for yield, 79.87% for fruit weight, 38.14% for fruit length, and 249.66% for the number of fruits per plant. On the other hand, Shrestha et al. (Reference Shrestha, Luitel and Kang2011) working with C. annuum found that the highest percentage of positive heterosis was obtained for fruit number (104.0%) and yield (141.2%), in the 5AVS7 × SP32 hybrid.
In addition, they obtained high and negative heterosis values for all the characters studied, which allowed inferring a wide range of genetic variability among the parents used in the crossing programme. Similar results were reported by Pech-May et al. (Reference Pech-May, Castañón-Nájera, Tun-Suárez, Mendoza-Elos, Mijangos-Cortés, Pérez-Gutiérrez and Latournerie-Moreno2010) in different sweet pepper crosses P3 × P5 (−65.7%), P2 × P7 (−54.3%), P1 × P6 (−42.4%), and P3 × P7 (−40.9%) for yield trait. These authors suggest that traits NFP and YP could be improved, using the hybridization method because it allows exploiting more efficiently the effects of nonadditive gene action, while for the variables fruit length and fruit diameter, low heterosis values are improved by the individual selection method. High and negative heterosis values were recorded for all the traits studied, which allows inferring the wide range of genetic variability between the parents used in the crossbreeding programme.
The Hb percentages obtained in our hybrids exceeded 100% for the NFP and YP variables. NFP was the yield component that contributed the most to this trait. This corroborates that reported by Pech-May et al. (Reference Pech-May, Castañón-Nájera, Tun-Suárez, Mendoza-Elos, Mijangos-Cortés, Pérez-Gutiérrez and Latournerie-Moreno2010), who also found that NFP made the greatest contribution to total fruit yield, supporting those genetic differences between parents promote hybrid vigour expression. From a practical perspective, (Hb) is a parameter of great interest for the commercial exploitation of hybrid vigour (Cabrera, Reference Cabrera2016). The same authors working with tomato crops found outstanding combinations exhibiting the best performance for the yield per plant variable compared to their superior parents. Prasad et al. (Reference Prasad, Reddy and Sadashiva2003) reported high and positive H and Hb values for chili (C. annuum) hybrids. Kumar and Lal (Reference Kumar and Lal2001) analysed heterosis and heterobeltiosis in hot pepper (C. annuum) hybrids and observed high Hb values for AFW, which allowed them to select high-yielding hybrids. Similarly, Shrestha et al. (Reference Shrestha, Luitel and Kang2011), working with sweet pepper (C. annuum) found high Hb values in the crosses 5AVS7 × SP32 (87.2%) and SP12 × SP38 (119.3%). In this work, there are other hybrids, although, to a lesser degree, they also showed Hb values above 100%, and could also be recommended for future crossbreeding programs. The wide diversity of the C. chinense species, combined with the high demand for Habanero peppers, has recently prompted different groups of scientists to work on the genetic improvement of this crop, to obtain varieties and hybrids that meet the expectations of the different markets (De Sousa and Maluf, Reference De Sousa and Maluf2003). Recent reports (Peña-Yam, Reference Peña-Yam2020; Ramírez, Reference Ramírez2020), show important advances that, in this sense, have been achieved. It is important to highlight, that the hybrid obtained through the RNJ-04 male tester, had a significant superiority over the rest of the crosses in terms of SCA, H, and Hb for most of the variables analysed, as well as for YP and its components. This will allow the effective selection of high-yielding F1 hybrids, with other important agronomic characteristics of great interest, for different markets that require fresh Habanero pepper for consumers, or as raw material to be processed.
The results obtained in this work, come to reaffirm the importance of generating F1 hybrids of Habanero pepper for fresh consumption, more productive, more uniform, that have less affected by the environment due to their heterozygous condition, and present greater productive potential than their parents. At the same time, that meets consumer expectations, due to the lack of improved varieties that have affected the slow development of this crop in the Yucatan region. Given the importance of hybridization as a breeding strategy, it has led current breeding programs to emphasize the development of these hybrids, due to their high yield potential based on the quality and productivity of this crop, taking advantage of the hybrid vigour that is generated, by combining in a single genotype, the favourable genes present in two or more different parents. The 16 hybrids obtained from Habanero pepper, come to form a genetic resource, which together with others already obtained, are the first to be achieved in the Peninsula from local varieties. With this, the independence of imported hybrid seeds is achieved, whose high cost makes it impossible for farmers in the area to acquire them (Peña-Yam et al., Reference Peña-Yam, Muñoz-Ramírez, Avilés-Viñas, Canto-Flick, Pérez-Pastrana, Guzmán-Antonio, Santana-Buzzy, Aguilera-Cauich and Mijangos-Cortés2019b).
The genetic improvement of plants allows obtaining varieties with characteristics of higher commercial and nutritional quality, greater resistance to abiotic and biotic adverse factors to crop, and higher yield. Through crossbreeding, different desired characteristics can be combined in the same individual, which, together with the selection process, will allow for achieving better and greater productive characteristics and a greater adaptation of the plants to the environment. The need to carry out a good selection process for Habanero pepper lines is becoming more important every day, in order to be used as more competitive F1 hybrid parents, where genetic-statistical parameters optimize this selection process. Recently, modern methods have emerged, that help to select the best characteristics and minimize the chances that the crops will be harmed by external factors, among which are molecular markers selection assisted, which combines very well with traditional breeding. Also, the induction of mutations when genetic variability is low; in vitro tissue culture, genetic engineering, and gene improvement, give the plant new beneficial characteristics such as resistance to diseases, pests, tolerance to herbicides, and stresses such as drought, frost, and high temperatures, and can contribute to changing the nutritional features in the fruits. Therefore, a sustainable and competitive varietal development is necessary to achieve new cultivars for different commercial purposes, with high yield potential, good climatic adaptation, and fruit quality, and that contemplates the production of F1 hybrids, due to the advantages that we offer (Rizwan et al., Reference Rizwan, Hussain, Shimelis, Hameed, Atif, Azhar and Asif2018).
This work offers information of great interest on the effects of GCA and specific (SCA) for the yield characters, and their components, of Habanero pepper genotypes, which allowed us to estimate the degree of dominance, heterosis, and heritability, with the purpose of identifying and evaluating heterotic patterns among the lines, developed from native germplasm of this crop, in Yucatan Peninsula. The varieties used in this study, have outstanding fruit quality characteristics in terms of flavour, aroma, pungency, and colour, that have been widely appreciated and they are different from what is produced in the rest of the country. For this reason, we consider that everything previously mentioned in this work is of great value, both theoretical and practical, considering that there is currently a lack of promising genetic materials to satisfy the growing demand for Habanero pepper, as well as a lack of basic information such as that obtained in this work for the successful development of improvement programs in this crop.
In conclusion, in the selection process, six of the ten parents stood out for having high and positive values for GCA, among them the parent NKA-11 obtained the highest values. The quantitative variables evaluated confirm an important genetic variability among the Habanero pepper genotypes studied, which allowed obtaining hybrids with superiority in SCA, H, and Hb, for most of the variables analysed, as well as for yield, with values higher than 4.29 Kg/Pta (85 tons/ha). The best-selected hybrids were NEK-03xRNJ-04, NBA-06xRNJ-04, AKN-07xRNJ-04, and RKI-10xRNJ-04, which show their great productive potential, a quality that would make them enter the national and international fresh market. Based on the GCA values of the parents and the heterosis of the progenies, it is concluded, that hybridization would be the most appropriate breeding method, to increase YP and NFP. These results will contribute to the development of future selection works in the genetic improvement of this crop.
Figure 1 shows the Habanero peppers F1 hybrids were selected for their high productive potential. (a) NEK-03 × RNJ-04, (b) NBA-06 × RNJ-04, (c) RKI-10 × RNJ-04 y (d) AKN-07 × RNJ-04.

Figure 1. Habanero peppers F1 hybrids selected for their high productive potential. (a) NEK-03 × RNJ-04, (b) NBA-06 × RNJ-04, (c) RKI-10 × RNJ-04 y (d) AKN-07 × RNJ-04.
Acknowledgements
We thank CONACYT for financing this research through the Basic Science and the Project of the call for National Problems 2016 with key 2016/2935, and for the Ph.D. scholarship granted to the student: CVU number 945005.
Author contributions
Rodriguez-Llanes Y, Iglesias-Andreu LG and Santana-Buzzy N conceptualization; Ku-Pech E data curation; Santana-Buzzy N funding acquisition; Rodriguez-Llanes Y, Guzmán-Antonio A, Mijangos-Cortés JO, Canto-Flick A, Avilés-Viña S, and Pijeira-Fernández G investigation; Pérez-Brito D, Mijangos-Cortés JO and Iglesias-Andreu LG methodology; Santana-Buzzy N project administration; Rodriguez-Llanes Y writing – original draft; Pérez-Brito D, Mijangos-Cortés JO, Iglesias-Andreu LG and Santana-Buzzy N writing – review & editing
Competing interest
The authors declare no competing interests.






