



Impact statement
Protecting and restoring nature depend on understanding the billions of decisions that people make. Such decisions range from simple acts like caring for wildlife in one’s garden to more complex decisions like what products to buy or which political candidate to support. These decisions are determined in part by direct experiences of, and relationships with, nature. These may affect nature directly (e.g., determining how much an individual disrupts wildlife habitats) or indirectly (e.g., affecting one’s thoughts and attitudes toward nature). Understanding how people’s relationships with nature differ, how they are changing and how they relate to people’s pro-nature attitudes and behaviours can help to reveal strategies that can benefit biodiversity. For example, people who feel more connected to nature are more likely to take action to help protect it. People’s relationships with nature might be improved, for example, by increasing the availability and accessibility of natural environments, and people’s inclination, ability and confidence to engage with nature. Such efforts have the potential to create a virtuous cycle of human–nature interactions, whereby increased engagement with nature leads to greater appreciation, enjoyment and desire to protect it. This is particularly important at a time when people’s relationships with nature are often declining.
Introduction
Scientific discussion of how to slow and reverse global biodiversity loss has concentrated far more on ecological solutions than on social change. This has been exemplified by papers published in the run-up to the Fifteenth Conference of the Parties (COP-15) to the UN Convention on Biological Diversity (CBD), where research focused strongly on how best to set targets and measure progress for conservation (e.g., Watson et al., Reference Watson, Keith, Strassburg, Venter, Williams and Nicholson2020; Obura et al., Reference Obura, Katerere, Mayet, Kaelo, Msweli, Mather, Harris, Louis, Kramer, Teferi, Samoilys, Lewis, Bennie, Kumah, Isaacs and Nantongo2021; Allan et al., Reference Allan, Possingham, Atkinson, Waldron, Di Marco, Butchart, Adams, Kissling, Worsdell, Sandbrook, Gibbon, Kumar, Mehta, Maron, Williams, Jones, Wintle, Reside and Watson2022; Leadley et al., Reference Leadley, Gonzalez, Obura, Krug, Londoño-Murcia, Millette, Radulovici, Rankovic, Shannon, Archer, Armah, Bax, Chaudhari, Costello, Dávalos, Roque, DeClerck, Dee, Essl, Ferrier, Genovesi, Guariguata, Hashimoto, Speranza, Isbell, Kok, Lavery, Leclère, Loyola, Lwasa, McGeoch, Mori, Nicholson, Ochoa, Öllerer, Polasky, Rondinini, Schroer, Selomane, Shen, Strassburg, Sumaila, Tittensor, Turak, Urbina, Vallejos, Vázquez-Domínguez, Verburg, Visconti, Woodley and Xu2022), the importance and maintenance of wilderness areas (e.g., Aycrigg et al., Reference Aycrigg, Mccarley, Belote and Martinuzzi2022; Pérez‐Hämmerle et al., Reference Pérez‐Hämmerle, Moon, Venegas‐Li, Maxwell, Simmonds, Venter, Garnett, Possingham and Watson2022), identifying priority areas for biodiversity conservation, and for expanding, and increasing the effectiveness of, protected area systems (e.g., Hanson et al., Reference Hanson, Rhodes, Butchart, Buchanan, Rondinini, Ficetola and Fuller2020; Ward et al., Reference Ward, Saura, Williams, Ramírez-Delgado, Arafeh-Dalmau, Allan, Venter, Dubois and Watson2020; Allan et al., Reference Allan, Possingham, Atkinson, Waldron, Di Marco, Butchart, Adams, Kissling, Worsdell, Sandbrook, Gibbon, Kumar, Mehta, Maron, Williams, Jones, Wintle, Reside and Watson2022; Brennan et al., Reference Brennan, Naidoo, Greenstreet, Mehrabi, Ramankutty and Kremen2022; Wauchope et al., Reference Wauchope, Jones, Geldmann, Simmons, Amano, Blanco, Fuller, Johnston, Langendoen, Mundkur, Nagy and Sutherland2022) and understanding the threats to, and recovery of, individual species (e.g., Grace et al., Reference Grace, Akçakaya, Bennett, Brooks, Heath, Hedges, Hilton‐Taylor, Hoffmann, Hochkirch, Jenkins, Keith, Long, Mallon, Meijaard, Milner‐Gulland, Rodriguez, Stephenson, Stuart, Young, Acebes, Alfaro‐Shigueto, Alvarez‐Clare, Andriantsimanarilafy, Arbetman, Azat, Bacchetta, Badola, Barcelos, Barreiros, Basak, Berger, Bhattacharyya, Bino, Borges, Boughton, Brockmann, Buckley, Burfield, Burton, Camacho‐Badani, Cano‐Alonso, Carmichael, Carrero, Carroll, Catsadorakis, Chapple, Chapron, Chowdhury, Claassens, Cogoni, Constantine, Craig, Cunningham, Dahal, Daltry, Das, Dasgupta, Davey, Davies, Develey, Elangovan, Fairclough, Febbraro, Fenu, Fernandes, Fernandez, Finucci, Földesi, Foley, Ford, Forstner, García, Garcia‐Sandoval, Gardner, Garibay‐Orijel, Gatan‐Balbas, Gauto, Ghazi, Godfrey, Gollock, González, Grant, Gray, Gregory, Grunsven, Gryzenhout, Guernsey, Gupta, Hagen, Hagen, Hall, Hallerman, Hare, Hart, Hartdegen, Harvey‐Brown, Hatfield, Hawke, Hermes, Hitchmough, Hoffmann, Howarth, Hudson, Hussain, Huveneers, Jacques, Jorgensen, Katdare, Katsis, Kaul, Kaunda‐Arara, Keith‐Diagne, Kraus, Lima, Lindeman, Linsky, Louis, Loy, Lughadha, Mangel, Marinari, Martin, Martinelli, McGowan, McInnes, Mendes, Millard, Mirande, Money, Monks, Morales, Mumu, Negrao, Nguyen, MdNH, Norbury, Nordmeyer, Norris, O’Brien, Oda, Orsenigo, Outerbridge, Pasachnik, Pérez‐Jiménez, Pike, Pilkington, Plumb, Portela, Prohaska, Quintana, Rakotondrasoa, Ranglack, Rankou, Rawat, Reardon, Rheingantz, Richter, Rivers, Rogers, Rosa, Rose, Royer, Ryan, Mitcheson, Salmon, Salvador, Samways, Sanjuan, Santos, Sasaki, Schutz, Scott, Scott, Serena, Sharma, Shuey, Silva, Simaika, Smith, JLY, Sultana, Talukdar, Tatayah, Thomas, Tringali, Trinh‐Dinh, Tuboi, Usmani, Vasco‐Palacios, Vié, Virens, Walker, Wallace, Waller, Wang, Wearn, Weerd, Weigmann, Willcox, Woinarski, Yong and Young2021; Mair et al., Reference Mair, Bennun, Brooks, Butchart, Bolam, Burgess, Ekstrom, Milner-Gulland, Hoffmann, Ma, Macfarlane, Raimondo, Rodrigues, Shen, Strassburg, Beatty, Mez-Creutzberg, Iribarrem, Irmadhiany, Lacerda, Mattos, Parakkasi, Tognelli, Bennett, Bryan, Carbone, Chaudhary, Eiselin, Fonseca, Galt, Geschke, Glew, Goedicke, Green, Gregory, Hill, Hole, Hughes, Hutton, Keijzer, Navarro, Lughadha, Plumptre, Puydarrieux, Possingham, Rankovic, Regan, Rondinini, Schneck, Siikamäki, Sendashonga, Seutin, Sinclair, Skowno, Soto-Navarro, Stuart, Temple, Vallier, Verones, Viana, Watson, Bezeng, Hm, Burfield, Clausnitzer, Clubbe, Cox, Freyhof, Gerber, Hilton-Taylor, Jenkins, Joolia, Joppa, Koh, Lacher, Langhammer, Long, Mallon, Pacifici, Polidoro, Pollock, Rivers, Roach, Rodríguez, Smart, Young, Hawkins and McGowan2021; Bolam et al., Reference Bolam, Ahumada, Akçakaya, Brooks, Elliott, Hoban, Mair, Mallon, McGowan, Raimondo, Rodríguez, Roe, Seddon, Shen, Stuart, Watson and Butchart2022). There is no doubt that these are all vitally important issues. However, the loss of biodiversity has been an outcome of many billions of decisions (with varying degrees of independence) by individual people. Such decisions, intentional and otherwise, include how people use and manage any natural resources that they have direct influence over (from domestic gardens and backyards to larger land and sea holdings), what resources and items they purchase as consumers, what organisations they encourage and assist (e.g., conservation NGOs), and which local, regional and national governmental policies and management interventions they support. These pathways, and how they can best be influenced, have long been studied within environmental sustainability. They have, however, attracted far less attention from the biodiversity conservation community.
This is not to say that behavioural decision making and social change have received no attention in the context of biodiversity conservation (Thomas-Walters et al., Reference Thomas-Walters, McCallum, Montgomery, Petros, Wan and Veríssimo2023). Interest has included such issues as managing demand for wildlife products (MacFarlane et al., Reference MacFarlane, Hurlstone, Ecker, Ferraro, van der Linden, Wan, Veríssimo, Burgess, Chen, Hall, Hollands and Sutherland2022), the promotion of farmers’ pro-environmental practices (Mastrangelo et al., Reference Mastrangelo, Gavin, Laterra, Linklater and Milfont2014), the application of ‘nudge theory’ (Nelson et al., Reference Nelson, Partelow and Schlüter2019), conservation messaging (Kidd et al., Reference Kidd, Garrard, Bekessy, Mills, Camilleri, Fidler, Fielding, Gordon, Gregg, Kusmanoff, Louis, Moon, Robinson, Selinske, Shanahan and Adams2019), the effectiveness of social marketing campaigns (Green et al., Reference Green, Crawford, Williamson and DeWan2019) and the influence of visual media on human–nature interactions (Silk et al., Reference Silk, Correia, Veríssimo, Verma and Crowley2021). Nonetheless, it does seem remarkable that, despite being raised at least a decade ago (e.g., St John et al., Reference St John, Edwards-Jones and Jones2010), it continues to be necessary for recent papers (including in high profile journals) to champion and highlight the role that the behavioural sciences, for example, could play in biodiversity conservation (e.g., Maynard et al., Reference Maynard, Monroe, Jacobson and Savage2020; Balmford et al., Reference Balmford, Bradbury, Bauer, Broad, Burgess, Burgman, Byerly, Clayton, Espelosin, Ferraro, Fisherj, Garnett, Jones, Marteau, Otieno, Polasky, Ricketts, Sandbrook, Sullivan-Wiley, Trevelyan, van der Linden, Veríssimo and Nielsen2021; Nielsen et al., Reference Nielsen, Marteau, Bauer, Bradbury, Broad, Burgess, Burgman, Byerly, Clayton, Espelosin, Ferraro, Fisher, Garnett, Jones, Otieno, Polasky, Ricketts, Trevelyan, van der, Veríssimo and Balmford2021).
A range of different viewpoints can help to understand how individual decisions are determined, the negative impacts on biodiversity and ways of reducing these (Clayton et al., Reference Clayton, Litchfield and Geller2013; Amel et al., Reference Amel, Manning, Scott and Koger2017; Reddy et al., Reference Reddy, Montambault, Masuda, Keenan, Butler, Fisher, Stanley and Gneezy2017; Ives et al., Reference Ives, Abson, von Wehrden, Dorninger, Klaniecki and Fischer2018). One is that of personalised ecology, which describes the set of direct interactions that each of us has with nature (Gaston et al., Reference Gaston, Soga, Duffy, Garrett, Gaston and Cox2018; Gaston, Reference Gaston2020; Soga and Gaston, Reference Soga and Gaston2022). Whilst the significance of such interactions, which are likely unique to each person in their composition, has long been recognised (e.g., Wilson, Reference Wilson1984; Kellert and Wilson, Reference Kellert and Wilson1993; Pyle, Reference Pyle1993; Stokes, Reference Stokes2006; Samways, Reference Samways2007), it has particularly come to the fore of recent (Clayton et al., Reference Clayton, Colléony, Conversy, Maclouf, Martin, Torres, Truong and Prévot2017; Soga and Gaston, Reference Soga and Gaston2022). This paper describes why personalised ecology provides a pertinent perspective by exploring the links with, and implications for, the future of biodiversity. Some of the issues discussed (e.g., biophilia, connection to nature, extinction of experience, shifting baselines) have been argued to be amongst the most vital for that future (e.g., Ehrlich and Kennedy, Reference Ehrlich and Kennedy2005; Kareiva, Reference Kareiva2008; Simaika and Samways, Reference Simaika and Samways2010). Given strong biases in the relevant research literature toward studies of culturally westernised societies, our considerations are similarly biased, although many may generalise more widely.
Personalised ecology
In the most fundamental sense, an individual’s personalised ecology describes all of their direct interactions with nature. This includes those with both micro- and macro-organisms. However, a narrower sense conception of personalised ecology, which is of more relevance in the present context, is the direct sensory interactions a person has with nature, predominantly through sight, sound, smell and touch. This is largely with macro-organisms. It is the focus on direct interactions which differentiates personalised ecology from broader considerations of ecosystem services (from which individual people frequently benefit without their necessarily interacting directly with the organisms providing such benefits; Gaston et al., Reference Gaston, Soga, Duffy, Garrett, Gaston and Cox2018).
Personalised ecology is concerned with interactions with nature. There has long been debate as to where the limits to what constitutes nature should lie and definitions can differ markedly, particularly across cultures and disciplines (e.g., Wohlwill, Reference Wohlwill and Wohlwill1983; Proctor, Reference Proctor1998; Wickson, Reference Wickson2008; Bratman et al., Reference Bratman, Hamilton and Daily2012; Hartig et al., Reference Hartig, Mitchell, de Vries and Frumkin2014; CBD, 2022). We use the same definition here as we have employed in other recent studies about human–nature interactions (e.g., Soga and Gaston, Reference Soga and Gaston2020, Reference Soga and Gaston2022), in which nature encompasses individual living organisms through to ecosystems, excluding those that are not self-sustaining. This enables a focus on essentially ‘wild’ organisms.
People’s direct interactions with some species and taxonomic groups have received much attention (e.g., sharks, snakes, bears; Chippaux, Reference Chippaux2017; Bombieri et al., Reference Bombieri, Delgado, Russo, Garrote, López-Bao, Fedriani and Penteriani2018; Gibbs, Reference Gibbs2021), often due to the perceived, potential or realised negative threat that they pose to people. On the whole, however, personalised ecologies remain poorly documented, and have been studied in relatively crude terms, for example, measuring the extent of greenspace in the vicinity of a person’s home or workplace, or the kind, frequency and duration of outdoor visits that they make (e.g., Shanahan et al., Reference Shanahan, Bush, Gaston, Lin, Dean, Barber and Fuller2016; Cox et al., Reference Cox, Plummer, Shanahan, Siriwardena, Fuller, Anderson, Hancock and Gaston2017b; White et al., Reference White, Alcock, Grellier, Wheeler, Hartig, Warber, Bone, Depledge and Fleming2019; Colley et al., Reference Colley, Irvine and Currie2022). Studies of human interactions with other species have almost invariably focused on the numbers and types of species that occur where an individual person lives or visits, rather than considering which ones, and in what numbers, an individual person actually encounters and experiences them (e.g., Fuller et al., Reference Fuller, Irvine, Devine-Wright, Warren and Gaston2007; Dallimer et al., Reference Dallimer, Irvine, Skinner, Davies, Rouquette, Armsworth, Maltby, Warren and Gaston2012; Methorst et al., Reference Methorst, Bonn, Marselle, Böhning-Gaese and Rehdanz2021).
Notwithstanding, it is apparent that personalised ecologies vary greatly amongst individual people, both within and between populations. On average, personalised ecologies will relate to the spatial variation of those components of biodiversity of which people tend to be more aware (e.g., larger organisms). In urbanised societies, and probably more widely, personalised ecologies can be very poor for many people. They are often also highly skewed such that the majority of nature interactions that do occur are experienced by only a small proportion of people (Cox et al., Reference Cox, Hudson, Shanahan, Fuller and Gaston2017a). In general, personalised ecologies are dependent on opportunity (e.g., the local presence and abundance of species), motivation (e.g., emotional affinity with nature) and capability (e.g., ability to see or hear particular species) (Dallimer et al., Reference Dallimer, Davies, Irvine, Maltby, Warren, Gaston and Armsworth2014; Soga and Gaston, Reference Soga and Gaston2022). These are in turn often related to socioeconomic circumstances. We return to these issues in more detail later.
Not only do personalised ecologies vary greatly between people, but an individual’s personalised ecology also varies across multiple time scales (Soga and Gaston, Reference Soga and Gaston2022). It changes through the day (often peaking when people are moving outdoors; Derks et al., Reference Derks, Giessen and Winkel2020), through the week (often peaking at weekends when people engage more in outdoor recreation; Veitch et al., Reference Veitch, Carver, Abbott, Giles-Corti, Timperio and Salmon2015), and through a person’s life course (often peaking both during childhood and during earlier periods of retirement; Hughes et al., Reference Hughes, Rogerson, Barton and Bragg2019). In much of the western world, personalised ecologies, especially those experienced by children, have also become progressively more limited across recent generations, a phenomenon referred to as extinction of experience (Pyle, Reference Pyle1993; Miller, Reference Miller2005; Soga and Gaston, Reference Soga and Gaston2016, Reference Soga and Gaston2023). On the other hand, international, and particularly intercontinental, travel has broadened (though not necessarily deepened) the personalised ecologies of a (typically small) minority of many human populations, allowing people to interact with species and ecosystems that are very different from those they would otherwise encounter. This is reflected most strongly through ecotourism.
Consequences for the future of biodiversity
People’s personalised ecologies have a wide array of consequences for the future of biodiversity. Most attention to positive direct interactions with nature has focused on the wellbeing benefits for people, with evidence of impacts on physical, psychological and social health (e.g., Keniger et al., Reference Keniger, Gaston, Irvine and Fuller2013; Hartig et al., Reference Hartig, Mitchell, de Vries and Frumkin2014; Bratman et al., Reference Bratman, Anderson, Berman, Cochran, Vries, Flanders, Folke, Frumkin, Gross, Hartig, Kahn, Kuo, Lawler, Levin, Lindahl, Meyer-Lindenberg, Mitchell, Ouyang, Roe, Scarlett, Smith, van den, Wheeler, White, Zheng and Daily2019; Marselle et al., Reference Marselle, Hartig, Cox, de, Knapp, Lindley, Triguero-Mas, Böhning-Gaese, Braubach, Cook, Vries, Heintz-Buschart, Hofmann, Irvine, Kabisch, Kolek, Kraemer, Markevych, Martens, Müller, Nieuwenhuijsen, Potts, Stadler, Walton, Warber and Bonn2021; Oh et al., Reference Oh, Zhang, Nghiem, Chang, Tan, Quazi, Shanahan, Lin, Gaston, Fuller and Carrasco2022). This has led to the development of a diversity of interventions to increase these benefits, focusing largely either on changing the environments in which people spend their time, or on changing their behaviour (Shanahan et al., Reference Shanahan, Astell-Burt, Barber, Brymer, Cox, Dean, Depledge, Fuller, Hartig, Irvine, Jones, Kikilus, Lovell, Mitchell, Niemelä, Nieuwenhuijsen, Pretty, Townsend, van Heezik, Warber and Gaston2019). The promotion of people–nature interactions for the purpose of improving human wellbeing does, of course, have the potential to benefit biodiversity directly (especially wild plants and animals living in urban areas), including through the creation and maintenance of accessible greenspaces that enable such interactions. However, there are a variety of other consequences of personalised ecologies, both positive and negative, that may have much greater importance for the future of biodiversity (Figure 1).
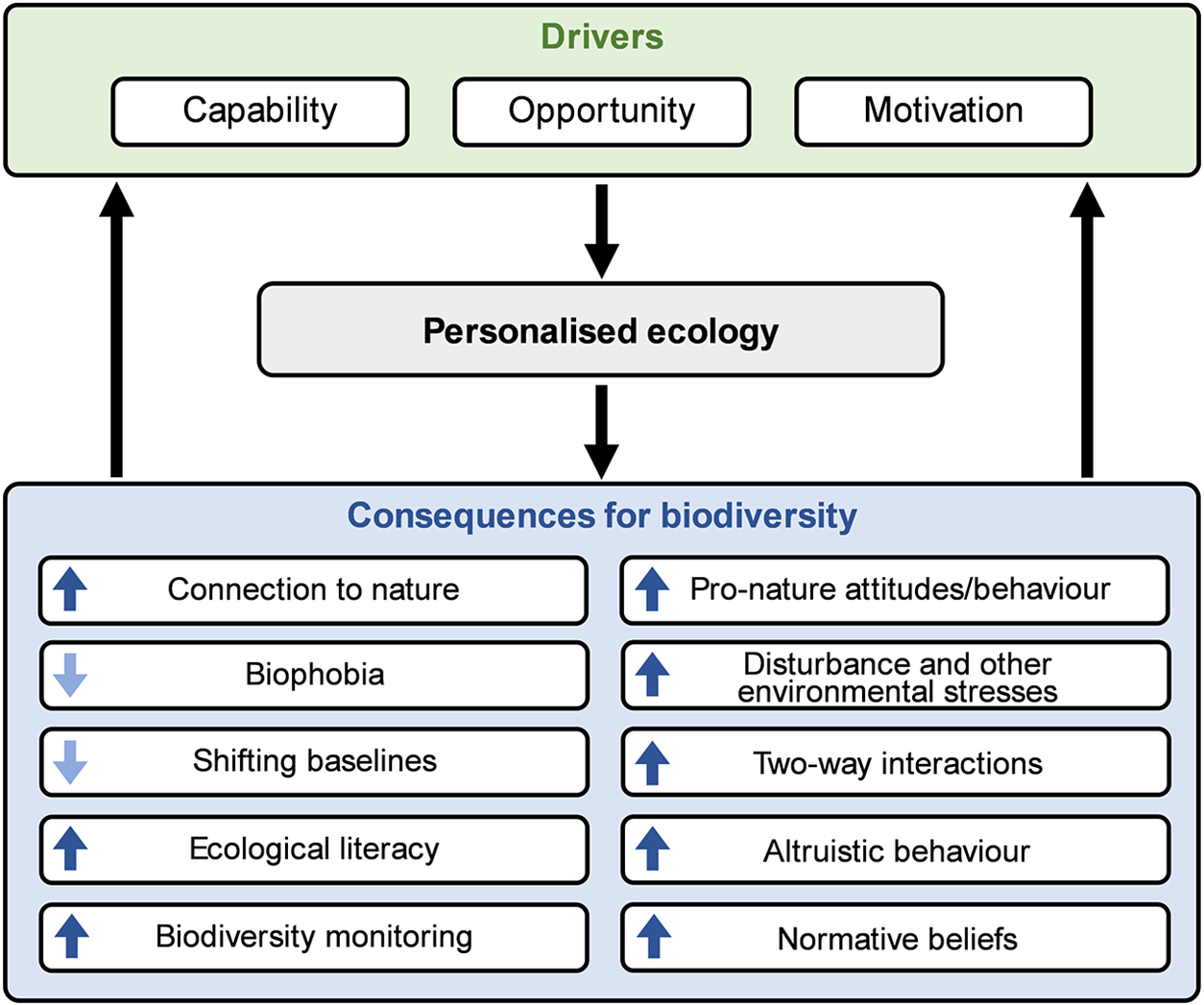
Figure 1. A conceptual diagram for understanding the drivers of personalised ecology and its consequences for biodiversity. There is likely a feedback loop in which the consequences of personalised ecology affect its drivers. In the consequences domain of Figure 1, each box contains an up or down arrow that denotes the direction of change in each factor or process caused by increased personalised ecology. For instance, the up arrow in the connection to nature box indicates that direct interactions with nature enhance one’s connection to nature.
(i) Connection to nature
People have a subjective, and perhaps innate, sense of connection with the natural world, sometimes known as biophilia (Wilson, Reference Wilson1984). Such nature connectedness varies dramatically amongst people and societies (Richardson et al., Reference Richardson, Hamlin, Elliott and White2022). It is increasingly seen as a core issue in human–nature relationships (Richardson et al., Reference Richardson, Dobson, Abson, Lumber, Hunt, Young and Moorhouse2020a), and meta-analyses have found that individuals with greater connection to nature have more pro-nature behaviours (Whitburn et al., Reference Whitburn, Linklater and Abrahamse2019; Barragan-Jason et al., Reference Barragan-Jason, de Mazancourt, Parmesan, Singer and Loreau2022). The strength of this connection to nature is thought to be enhanced in individuals with a richer and deeper personalised ecology and, conversely, to be weakened in those whose personalised ecology is poorer (Richardson et al., Reference Richardson, Passmore, Barbett, Lumber, Thomas and Hunt2020b; Mikołajczak et al., Reference Mikołajczak, Lees, Barlow, Sinclair, Trindade de Almeida, Souza and Parry2021; Li et al., Reference Li, Zhai, Chang, Merrill, Browning and Sullivan2022; Lim et al., Reference Lim, Sing, Chong, Jaturas, Dong, Lee, Tao, Le, Bonebrake, TPN, Chu, Brandon-Mong, Kong, Soga and Wilson2022). Indeed, whilst Wilson (Reference Wilson1984) defined biophilia as ‘the innate tendency to focus on life and lifelike processes’, in subsequent writings he emphasised that it is a complex set of learned behaviours, that is, a disposition that is reinforced, amplified and expressed through human culture (Wilson, Reference Kellert and Wilson1993). This is now supported by empirical evidence (Figure 2A; Collado et al., Reference Collado, Staats and Corraliza2013; Vanderstock et al., Reference Vanderstock, Grandi-Nagashiro, Kudo, Latty, Nakamura, White and Soga2022; Wu et al., Reference Wu, Ji and Jin2023). Connection to nature has also been found to mediate the link between personalised ecologies and pro-nature behaviours (Liu et al., Reference Liu, Cleary, Fielding, Murray and Roiko2022).
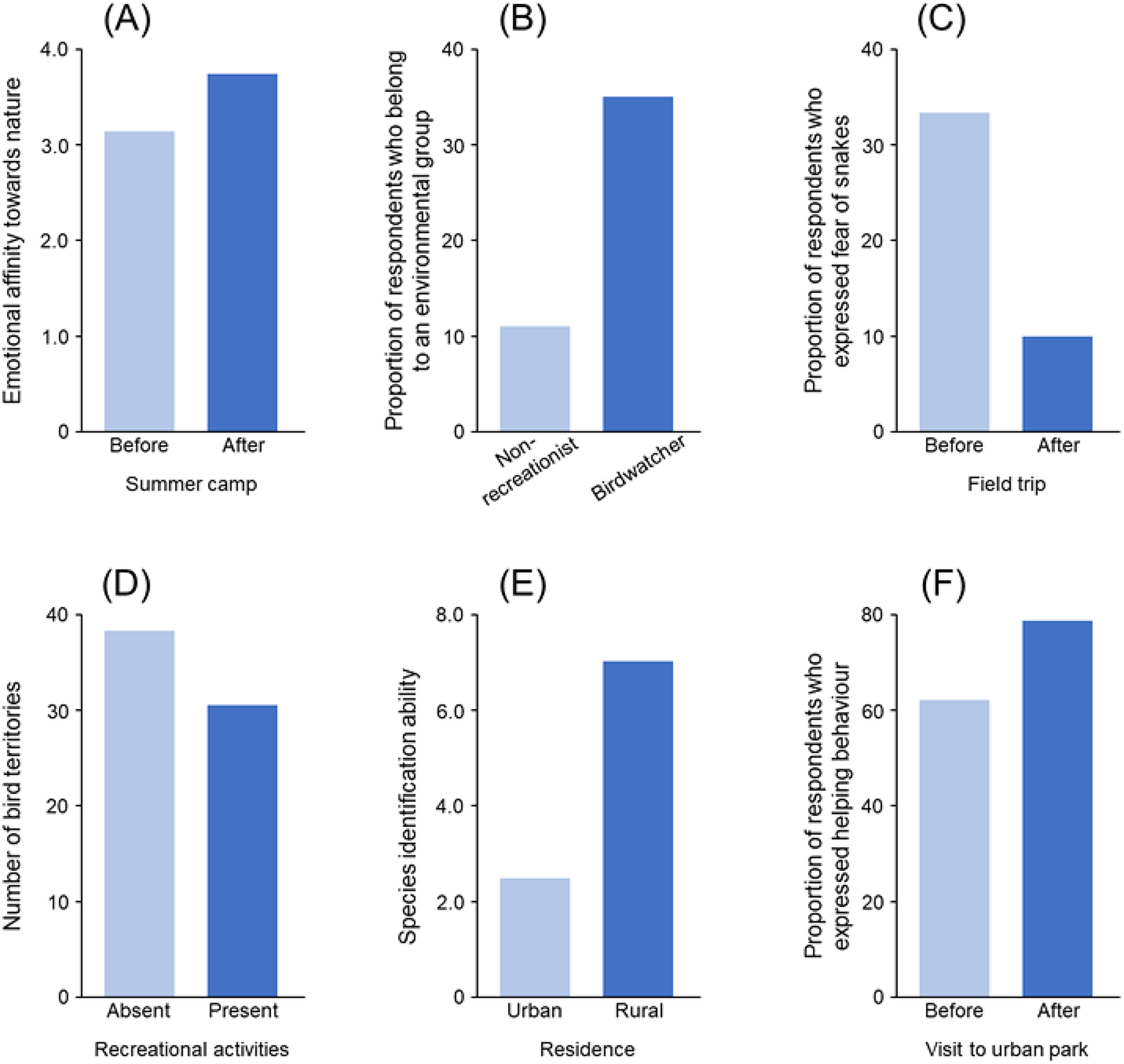
Figure 2. Empirical evidence suggesting several possible impacts of increased personalised ecology on biodiversity (A: connection to nature; B: pro-nature attitudes/behaviour; C: biophobia; D: disturbance of wildlife; E: ecological literacy; F: altruistic behaviour). Plots show: (A) changes in emotional connection to nature (measured by the Emotional Affinity toward Nature scale) before and after participating in a nature-based recreational program (summer camps) (Collado et al., Reference Collado, Staats and Corraliza2013); (B) differences in the likelihood of engaging in a pro-nature behaviour between birdwatchers and those who do not use nature for recreational purposes (non-recreationist) (Cooper et al., Reference Cooper, Larson, Dayer, Stedman and Decker2015); (C) changes in the proportion of people exhibiting fear of snakes before and after participating in a field trip (Ballouard et al., Reference Ballouard, Provost, Barré and Bonnet2012); (D) differences in the number of bird territories between sites with and without recreational activities (Bötsch et al., Reference Bötsch, Tablado and Jenni2017); (E) differences in species identification ability between people who live in urban versus rural areas (Bashan et al., Reference Bashan, Colléony and Shwartz2021); and (F) changes in the proportion of people exhibiting helping behaviour before and after experience of an urban green park (Guéguen and Stefan, 2014).
(ii) Pro-nature attitudes and behaviours
A key question relating to personalised ecologies is whether they influence a person’s pro-nature attitudes and behaviours, which has recently been termed the nature benefit hypothesis (Soga and Gaston, Reference Soga and Gaston2022). Several studies have documented positive relationships between levels of nature experience and pro-environmental attitudes and behaviours (Figure 2B; e.g., Wells and Lekies, Reference Wells and Lekies2006; Zelenski et al., Reference Zelenski, Dopko and Capaldi2015; Broom, Reference Broom2017; Rosa et al., Reference Rosa, Profice and Collado2018; Dean et al., Reference Dean, Barnett, Wilson and Turrell2019; Alcock et al., Reference Alcock, White, Pahl, Duarte-Davidson and Fleming2020; Martin et al., Reference Martin, White, Hunt, Richardson, Pahl and Burt2020; Liu and Chen, Reference Liu and Chen2021; Ngo et al., Reference Ngo, Hosaka and Numata2022). A smaller number have tested for and documented positive relationships for more explicitly pro-nature (a subset of pro-environmental) attitudes and behaviours (Cooper et al., Reference Cooper, Larson, Dayer, Stedman and Decker2015; Soga et al., Reference Soga, Gaston, Yamaura, Kurisu and Hanaki2016; Prévot et al., Reference Prévot, Cheval, Raymond and Cosquer2018).
(iii) Biophobia
Whilst reduced positive interactions with nature may weaken support for biodiversity conservation, there is an additional concern that these reductions may strengthen antagonism toward such interactions, that is, a negative feedback loop whereby the less that people interact with nature the less they want to do so. This could occur if the loss of positive nature interactions resulted in an increase in wariness or phobia toward nature, that is, biophobia (Ulrich, Reference Ulrich, Kellert and Wilson1993). Indeed, there is evidence that extinction of experience is associated with an increase in biophobia, including due to its associated loss of knowledge about nature (e.g., ability to identify species; Figure 2C; Ballouard et al., Reference Ballouard, Provost, Barré and Bonnet2012; Silva and Minor, Reference Silva and Minor2017; Ngo et al., Reference Ngo, Hosaka and Numata2019; Soga et al., Reference Soga, Evans, Yamanoi, Fukano, Tsuchiya, Koyanagi and Kanai2020; Fukano and Soga, Reference Fukano and Soga2021; Sugiyama et al., Reference Sugiyama, Hosaka, Takagi and Numata2021).
Elevated biophobia can have a wide range of detrimental impacts on the future of biodiversity. Fear emotions impose a significant psychological cost for humans, and thus, increased biophobia can reduce the willingness of local people to coexist with wild animals, particularly, those regarded as dangerous or harmful (e.g., wolves, bears, large cats). Biophobia can therefore decrease public acceptance of certain policies and actions to conserve and restore these organisms (e.g., reintroduction). Biophobia also often results in an increase in persecution of wild organisms, which can negatively impact biodiversity more directly (Pandey et al., Reference Pandey, Subedi Pandey, Devkota and Goode2016; Rocha et al., Reference Rocha, Aziz, Brook, Carvalho, Cooper-Bohannon, Frick, Huang, Kingston, López-Baucells, Maas, Mathews, Medellin, Olival, Peel, Plowright, Razgour, Rebelo, Rodrigues, Rossiter, Russo, Straka, Teeling, Treuer, Voigt and Webala2021). If increasing urbanisation of the human population, and general decline of biodiversity, result in increased biophobia, the impacts on the future of biodiversity could be severe.
(iv) Negative impacts of nature engagement on biodiversity
Increased human–nature interactions may lead to negative impacts on biodiversity in several ways. This includes, for example, disturbance of wildlife during recreational activities (Figure 2D; Larson et al., Reference Larson, Reed, Merenlender and Crooks2016; Bötsch et al., Reference Bötsch, Tablado and Jenni2017), loss of predator avoidance behaviour (Geffroy et al., Reference Geffroy, Samia, Bessa and Blumstein2015), the unintentional transport of organisms between sites (including both native and non-native species) (Hodkinson and Thompson, Reference Hodkinson and Thompson1997), changes to understorey vegetation (Erfanian et al., Reference Erfanian, Alatalo and Ejtehadi2021), increased chemical pollutants (e.g., negative impacts of sunscreen use on coral reefs; Danovaro et al., Reference Danovaro, Bongiorni, Corinaldesi, Giovannelli, Damiani, Astolfi, Grec and Pusceddu2008), increased litter and more frequent fires. Better connection to nature might therefore have negative consequences if it means that more people are going out and disturbing or damaging flora and fauna in sensitive areas.
(v) Two-way interactions
There is evidence of an asymmetry in people’s beliefs, whereby they commonly hold that human impacts on the natural environment are greater than the impacts of the natural environment on people (e.g., Coley et al., Reference Coley, Betz, Helmuth, Ellenbogen, Scyphers and Adams2021). This can weaken the role of self, family or human benefits in support for pro-nature behaviours, and is clearly at odds with the utter dependence of humanity on ecosystem services (IPBES, Reference Brondizio, Settele, Díaz and Ngo2019).
(vi) Shifting baselines
The personalised ecologies that people experience, particularly earlier in life, can have a profound impact on what one regards as ‘normal’ and ‘sound’. Faced with declines in the state of nature, this can result in a progressive ‘ratcheting down’ or shifting of baselines (Pauly, Reference Pauly1995; Soga and Gaston, Reference Soga and Gaston2018). People may, therefore, become more accepting of a much-depleted biodiversity, because the extent of the departure from a natural situation is poorly understood (Jones et al., Reference Jones, Turvey, Massimino and Papworth2020). Shifting baselines can have many negative impacts on biodiversity conservation as they may lead to an increased tolerance for the progressive degradation of ecosystems, changes in people’s expectations as to the state of nature that is worth protecting or restoring, and subsequently the establishment of less ambitious targets and goals for nature conservation (Soga and Gaston, Reference Soga and Gaston2018).
(vii) Ecological literacy
Reduced positive interactions with nature can weaken people’s knowledge about local ecosystems (Figure 2E; Bashan et al., Reference Bashan, Colléony and Shwartz2021). This is often called ecological literacy, or eco-literacy (Pilgrim et al., Reference Pilgrim, Smith and Pretty2007). It includes, for example, identification skills of local fauna and flora (Bashan et al., Reference Bashan, Colléony and Shwartz2021), ethnobotanical knowledge (e.g., traditional use of edible/medicinal plants; Okui et al., Reference Okui, Sawada and Yoshida2021), and traditional management practices of local ecosystems (Tsuchiya et al., Reference Tsuchiya, Aoyagi, Okuro and Takeuchi2014). Maintenance of such knowledge is fundamental for the continued support of local conservation efforts and the capacity of local communities to self-manage natural resources sustainably. Declines in local ecological knowledge can therefore have negative impacts on the conservation of biodiversity.
(viii) Altruistic behaviour
There is evidence that exposure to nature (e.g., viewing greenery) can contribute to enhancing altruism in humans, which has recently been termed the nature and sustainability hypothesis (Soga and Gaston, Reference Soga and Gaston2022). This includes various behaviours, including the reduction of impulsive and selfish decision making and the promotion of sustainable, cooperative and helping behaviour (Figure 2F; Van der Wal et al., Reference Van der Wal, Schade, Krabbendam and Van Vugt2013; Zelenski et al., Reference Zelenski, Dopko and Capaldi2015; Guéguen and Stefan, Reference Guéguen and Stefan2016). Increased altruistic decision making and behaviour can have a wide range of positive outcomes for biodiversity as those actions can contribute, either directly or indirectly, to the conservation and restoration of wild plants and animals.
(ix) Biodiversity data collection
Increased nature interactions may, in some cases, contribute to an increased amount of biodiversity data coming from citizens (Schuttler et al., Reference Schuttler, Sorensen, Jordan, Cooper and Shwartz2018). For example, it has been suggested that increased use of urban greenspaces during the COVID-19 pandemic resulted in increased numbers of wildlife observations submitted to citizen science projects (e.g., Hochachka et al., Reference Hochachka, Alonso, Gutiérrez-Expósito, Miller and Johnston2021). Citizen science data can offer a valuable source of species occurrence records and be used to generate species-level information for broad-scale biodiversity mapping and monitoring.
(x) Normative beliefs
Normative beliefs are the perceptions of what are socially typical or acceptable attitudes and behaviours. For a particular person, both their personalised ecology, and the actual and perceived personalised ecologies of those around them, will shape their normative beliefs. Normative beliefs are often a strong predictor of people’s attitudes and behaviours (Armitage and Conner, Reference Armitage and Conner2010), including those relating to biodiversity (van Riper et al., Reference Van Riper, Browning, Becker, Stewart, Suski, Browning and Golebie2019). Social norms and normative beliefs may influence attitudes and connection to nature (Oh et al., Reference Oh, Fielding, Nghiem, Chang, Carrasco and Fuller2021), or influence behaviour directly in the form of cultural taboos toward the exploitation of particular species, areas and natural resources (Jones et al., Reference Jones, Andriamarovololona and Hockley2008), or more subtly through a person’s propensity to engage in pro-nature behaviours, such as participation in urban greenspace management (Marshall et al., Reference Marshall, Grose and Williams2020).
Acting in combination
Of course, these 10 consequences of personalised ecologies do not act independently, but likely generate a synergistic effect on biodiversity conservation. For example, if people obtain eco-literacy through enhanced personalised ecologies, they may use natural environments in a way that does less harm to those environments (e.g., maintaining appropriate distances from wildlife). Likewise, those with greater species identification ability can provide more accurate and reliable data on biodiversity. Further, increased connection to nature is known to act as a protective factor against biophobia (Zsido et al., Reference Zsido, Coelho and Polák2022). However, there may equally be negative synergies between some of these consequences. For example, shifting baselines are likely to interact with normative beliefs because social norms (perceived or actual) provide a self-sustaining mechanism for maintaining poor personalised ecologies. This may result in a negative, self-reinforcing feedback loop, making it difficult to reverse historic declines in people’s personalised ecologies.
Strengthening personalised ecologies
If better developed personalised ecologies generate positive outcomes for biodiversity conservation, as described above, then strengthening those ecologies may be critically important for the future of biodiversity. People’s personalised ecologies can usefully be regarded as being shaped by three broad sets of factors: capability, opportunity and motivation (as distinguished by the COM-B model; Michie et al., Reference Michie, Van Stralen and West2011). Each of these provides a unique set of opportunities and approaches that could be targeted to strengthen personalised ecologies.
(i) Capability
Capability is an individual’s capacity to engage in interactions with nature. It has two components, physical capability and psychological capability. Physical capability includes the ease with which one can move around and the extent to which one has sufficient sensory abilities to detect particular species, for example, being able to see birds or hear birdsong. Psychological capability includes knowledge, skills, stamina and confidence. The component that has attracted the most attention is skills such as the ability to recognise particular species (Figure 3A; Lindemann-Matthies, Reference Lindemann-Matthies2002).
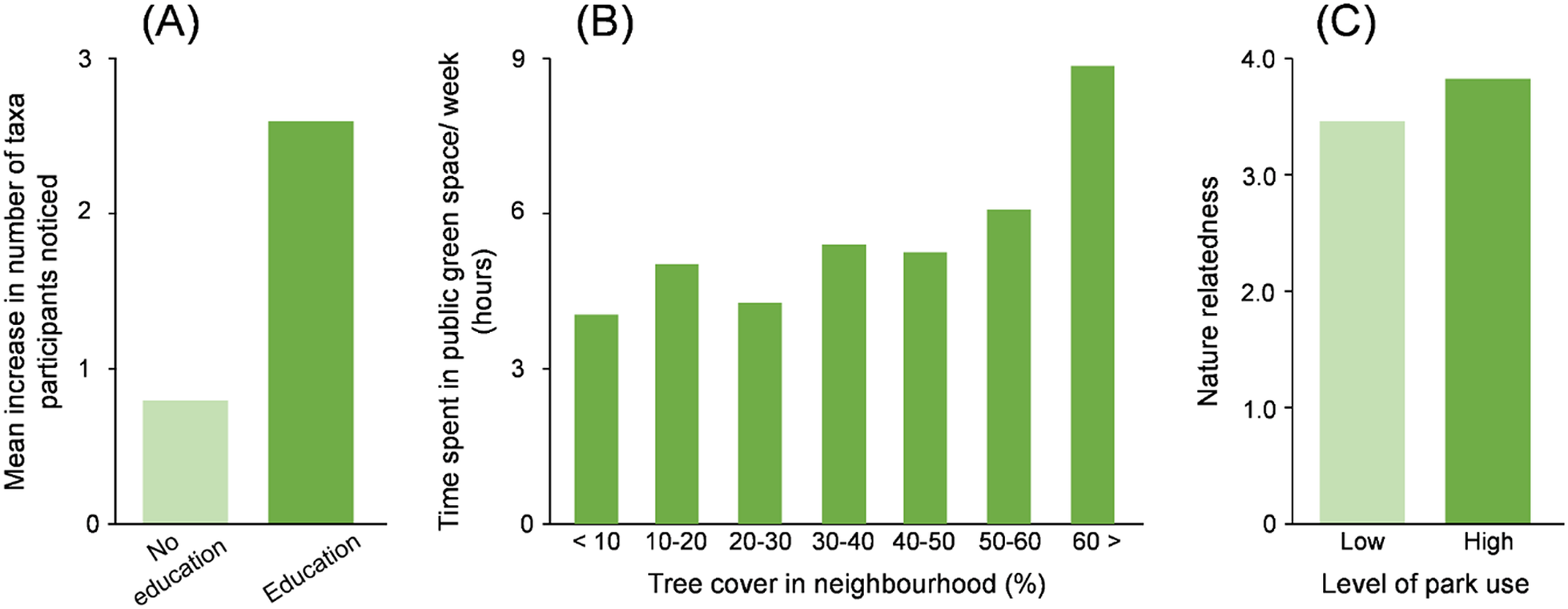
Figure 3. Empirical evidence demonstrating the role of (A) capability, (B) opportunity and (C) motivation in determining personalised ecology. Plots show: (A) effects of an educational program aimed at increasing children’s species identification ability on the number of animal and plant taxa children noticed on the way to school (Lindemann-Matthies, Reference Lindemann-Matthies2002); (B) association between neighbourhood tree cover and time spent in public greenspace (Shanahan et al., Reference Shanahan, Cox, Fuller, Hancock, Lin, Anderson, Bush and Gaston2017); and (C) differences between park users (based on time spent in parks) in terms of their emotional connection to nature (measured by the Nature Relatedness scale; Lin et al., Reference Lin, Fuller, Bush, Gaston and Shanahan2014).
Arguably, biodiversity conservation has been heavily fixated on improving a rather narrow conception of capability – assuming that education about the nature around you will improve your ability to access it, and willingness to protect it (Thomas-Walters et al., Reference Thomas-Walters, McCallum, Montgomery, Petros, Wan and Veríssimo2023). This is despite various studies finding that education and knowledge, by themselves, are relatively poor predictors of connection to nature (e.g., Barragan-Jason et al., Reference Barragan-Jason, de Mazancourt, Parmesan, Singer and Loreau2022) and pro-nature behaviours (e.g., Knapp et al., Reference Knapp, Phillips, Clements, Shaw and Osborne2021).
Capability might be improved by (i) improving ways for less physically able people to interact with nature (e.g., via views from windows, improved access to greenspaces); (ii) equipment that enables people to overcome or reduce sensory limitations in interacting with nature (e.g., vision and acoustic systems); (iii) accessible tools and learning that help improve psychological capability; and (iv) guides (particularly people rather than signage) who can facilitate and explain nature interactions to visitors to sites.
(ii) Opportunity
Opportunity is all of the factors in a person’s environment that make interactions with nature possible. It has two components, physical opportunity and social opportunity. Physical opportunity includes the availability of nature in a person’s environment with which they can interact (Figure 3B; Shanahan et al., Reference Shanahan, Cox, Fuller, Hancock, Lin, Anderson, Bush and Gaston2017). Social opportunity includes family values, social norms and public safety. Attention has particularly focused on the role of physical opportunity in personalised ecology, and the extent to which people have adequate or appropriate access to nature in their immediate neighbourhood or more widely. Indeed, some organisations have established targets for the availability of local greenspace, such as at least 0.5 ha within 200 m, 2 ha within 300 m and 10 ha within 1 km, all within a 15 min walk from home (Natural England, 2022). Social opportunity, on the other hand, has received much less attention in discussions on how to promote people’s use of nature, except for some particular cases such as children’s use of local nature (Button et al., Reference Button, Tillmann and Gilliland2020). However, recent studies suggest that the influence of social opportunity on personalised ecologies is comparable to – and sometimes stronger than – that of physical opportunity (e.g., Soga et al., Reference Soga, Yamanoi, Tsuchiya, Koyanagi and Kanai2018; Van Truong et al., Reference Van Truong, Nakabayashi and Hosaka2022).
Opportunity might be improved by: (i) improving the availability and accessibility of local nature, including in the vicinity of both home and work places; (ii) improving the ability of nature to move amongst greenspaces, influencing both species’ population sizes and the potential for human–nature interactions; (iii) physically ‘greening’ buildings and their immediate surroundings, through green roofs and walls, gardens, etc.; (iv) improving transport systems to enable better access to nature sites; (v) changing and challenging values, social norms and normative beliefs around nature interactions (e.g., through community engagement, community champions and role models); (vi) improving safety of the local environment (e.g., improving road safety, reducing crime, controlling the abundance of wild animals that can have severe negative health impacts on people); and (vii) providing more dedicated time for nature interactions (e.g., built into work/school schedules).
(iii) Motivation
This is the set of brain processes that energise and direct behaviour. Its two components are: automatic motivation and reflective motivation. Automatic motivations are unconscious responses, such as emotional reactions, whilst reflective motivations are more cognitive and purposeful, such as intentions (Figure 3C; Lin et al., Reference Lin, Fuller, Bush, Gaston and Shanahan2014). In the field of human–nature interactions, the importance of motivation (particularly automatic motivation) in promoting personalised ecologies has long been recognised, and indeed studies show that it is often the most impactful factor in predicting the quantity and quality of those interactions (e.g., Lin et al., Reference Lin, Fuller, Bush, Gaston and Shanahan2014; Soga and Akasaka, Reference Soga and Akasaka2019). Of course, motivation is likely to be improved by enhanced personalised ecologies (see earlier discussion on biophilia), implying that there exists a bidirectional relationship between motivation and personalised ecology.
Motivation might be improved by: (i) green social prescribing, which can provide an incentive for reflective motivation and intentions to interact with nature; (ii) nature-based educational programs in educational institutions (e.g., schools, museums) that can help to increase connection to nature, and therefore automatic motivation; and (iii) nature-oriented television and internet programs (e.g., nature documentaries), and social media that promote people’s desire to experience nature.
Strategies to improve opportunity, motivation and capability do not work independently but are interrelated in many ways. For example, providing nature-based education in schools can help to increase all of the three drivers (capability: ability to notice wildlife; opportunity: ensuring time to interact with nature; motivation: nature connectedness). Improving emotional connection to nature, through recreational and educational programs, is also known to be closely related to enhanced psychological wellbeing (Pirchio et al., Reference Pirchio, Passiatore, Panno, Cipparone and Carrus2021), suggesting that it may help to increase psychological capability.
Conclusions
Many factors shape people’s behavioural decisions, small or large, which collectively determine the future of biodiversity. People’s personalised ecologies are a central factor that may act directly (impacting nature during people’s interactions) or indirectly (influencing mediating factors such as attitudes, nature connectedness and normative beliefs). This raises the potential of a virtuous cycle whereby improving personalised ecologies encourages demand for improved biodiversity, at a time when a high proportion of the global population’s interactions with nature are extremely constrained, and becoming poorer.
Open peer review
To view the open peer review materials for this article, please visit http://doi.org/10.1017/ext.2023.15.
Data availability statement
Data availability is not applicable to this article as no new data were created or analysed in this study.
Acknowledgements
We are grateful to three anonymous reviewers for their thoughtful comments.
Author contribution
Conception and design of work: K.J.G., B.B.P. and M.S.; Drafting and revising: K.J.G., B.B.P. and M.S.
Financial support
K.J.G. and B.B.P. were supported by the Natural Environment Research Council funded ‘Renewing biodiversity through a people-in-nature approach (RENEW)’ project (NE/W004941/1). M.S. was supported by the Japan Society for the Promotion of Science (Grant Nos. 20H04375 and 23H03583).
Competing interest
The authors declare no competing interest exists.




 Open access
Open access
Comments
Please find herewith our invited contribution to ‘Extinction’, addressing as agreed the relations between personalised ecology and the future of biodiversity. We trust that you will find that this is appropriate. Please do not hesitate to contact us if you have any queries.