



INTRODUCTION
The Hoxnian interglacial period is of particular interest for its archaeological evidence of human occupation in Britain and its correlation with Marine Isotope Stage (MIS) 11c, 426.6 to 395.9 ka (Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008; Rodrigues et al., Reference Rodrigues, Voelker, Grimalt, Abrantes and Naughton2011; Candy et al., Reference Candy, Tye, Coxon, Hardiman, Matthews and Palmer2021), a good past analog for the present warm period (Loutre and Berger, Reference Loutre and Berger2003; Candy et al., Reference Candy, Schreve, Sherriff and Tye2014). Long, continuous, deep-marine sequences offer complete records of MIS 11 (duration ca. 420 to 360 ka; Loutre and Berger, Reference Loutre and Berger2003) that include not only the interglacial (MIS 11c) but also post-interglacial intervals (MIS 11b, MIS 11a). In Britain, the Hoxnian interglacial period is well recorded, sometimes together with other MIS 11 intervals, in sequences that are predominantly lacustrine at Hoxne (stratotype), Marks Tey (parastratotype) (West, Reference West1956; Turner, Reference Turner1970; Thomas, Reference Thomas2001), Barnham (Ashton et al., Reference Ashton, Lewis and Parfitt1998, Reference Ashton, Lewis, Parfitt, Davis and Stringer2016), and Elveden (Ashton et al., Reference Ashton, Lewis, Parfitt, Candy, Keen, Kemp, Penkman, Thomas, Whittaker and White2005); terrestrial at West Stow (Preece et al., Reference Preece, Parfitt, Bridgland, Lewis, Rowe and Atkinson2007); and fluvial at Southfleet Road, Ebbsfleet (Wenban-Smith, Reference Wenban-Smith2013), Swanscombe (White et al., Reference White, Preece and Whittaker2013), Quinton (Coope and Kenward, Reference Coope and Kenward2007), and Woodston (Horton et al., Reference Horton, Keen, Field, Robinson, Coope, Currant, Graham, Green and Phillips1992) (Fig. 1). At Hoxne (Fig. 1), the Anglian late glacial is represented by Stratum F and the Hoxnian interglacial (pollen zones HoI to HoIIIa) by Strata E and D. The lacustrine sediments of Strata F (clay with chalk pebbles) and E (unstratified clay) are capped by peat (Stratum D) representing the encroachment of terrestrial vegetation. The absence of pollen zones HoIIIb and HoIV and the occurrence of reworked plant material in Stratum C indicate a substantial hiatus between Stratum D and a return to lacustrine deposition (laminated sands, silts, and clays) under cold climatic (stadial) conditions in Stratum C (the “Arctic Bed”) (Reid, in Evans et al., Reference Evans, Morse, Reid, Ridley and Ridley1896; West, Reference West1956; Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008). Archaeological evidence of human occupation is confined to the succeeding interstadial deposits: fluvial sediments (chalky gravels, sands, silts, and clays) of Stratum B and overlying alluvial deposits (gravels, sands, and sandy clays) and solifluction gravels of Stratum A (West, Reference West1956; Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008). Stratum B has yielded a temperate-climate vertebrate assemblage of mammals, birds, amphibians, and fish (the last including rudd, Scardinius erythrophthalmus, indicative of warm summer water temperatures); the lower part of Stratum A contains a similarly temperate assemblage that may, however, be reworked from Stratum B, and the upper part of Stratum A shows evidence of cold (periglacial) conditions. Ashton et al. (Reference Ashton, Lewis, Parfitt, Penkman and Coope2008) compared the Hoxne sequence with palynological records from key European sites and a marine core off the northwest coast of the Iberian Peninsula, as well as other proxy records from Lake Baikal sediments and an Antarctic ice core, and tentatively suggested that Strata C–A at Hoxne might be correlated with one of the cold/warm cycles in the later part of MIS 11 (implying that the hiatus between Strata D and C lasted ca. 20,000 yr). They argued that since the archaeology at Hoxne is associated with the later interstade, rather than the earlier interglacial period, humans were living in a boreal forest environment, with winters significantly colder than during the fully temperate interglacial. This leads to questions about the possible technologies (clothing, shelters, fire) or physiological adaptations that enabled humans to survive harsher winters (Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008; Ashton and Lewis, Reference Ashton and Lewis2012; Hosfield, Reference Hosfield2016).

Figure 1. (A) Location of British MIS 11 sites mentioned in the text and Table 1; (B) summary succession of MIS 11 deposits at Hoxne; (C) schematic cross section of the Hoxne lake basin; (D) map of the Hoxne lake basin showing sampling locations referred to in the text and Supplementary Material. C and D modified from Ashton et al. (Reference Ashton, Lewis, Parfitt, Penkman and Coope2008), with data from West (Reference West1956).
Quantitative palaeoclimate reconstructions are of key importance in informing such discussion, but so far the available palaeotemperature reconstructions for Hoxne have been limited to those based on beetles for Strata F, D, and C (Coope, Reference Coope, Singer, Gladfelter and Wymer1993; Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008), with the addition of an ostracod-based reconstruction of Stratum C palaeotemperatures by Benardout (Reference Benardout2015) that disagrees with the beetle-based reconstruction. Quantitative palaeotemperature reconstructions for the Hoxnian interglacial and post-interglacial intervals of MIS 11 have been published for five other British sites, each based on the application of a single proxy method (Table 1).
Table 1. Summary of published quantitative palaeotemperature reconstructions for British MIS 11 sites.

a BMCR, beetle mutual climatic range; MOTR, mutual ostracod temperature range.
In studies of British Pleistocene sites, single-proxy quantitative reconstructions have often been accepted uncritically; however, any single proxy method on its own is likely to produce a plausible (though not necessarily accurate) result, and there is a pressing need to test, validate, and refine reconstructions by means of multiproxy applications. If the results of two or more proxy methods are correct, then they should have overlapping ranges; in such a case, the mutual overlap of the reconstructed palaeotemperatures can be used to identify a narrower range within which all the proxies could have coexisted. If the results from two or more proxies do not overlap, the results must be treated with caution (if not simply rejected) and the reasons for the discrepancy explored. Holmes et al. (Reference Holmes, Atkinson, Darbyshire, Horne, Joordens, Roberts, Sinka and Whittaker2010) cross-checked and combined ostracod and herpetile palaeotemperature reconstructions to validate and refine mean July, January, and annual air temperature ranges for MIS 13 at Boxgrove. At Whittlesey, combinations of beetle and ostracod mutual climatic range methods with vertebrate indicator taxa evidence have yielded robust palaeotemperature reconstructions for the MIS 7–6 transition (Langford et al., Reference Langford, Boreham, Briant, Coope, Horne, Schreve, Whittaker and Whitehouse2014a), MIS 7 (Langford et al., Reference Langford, Boreham, Coope, Fletcher, Horne, Keen, Mighall, Penkman, Schreve and Whittaker2014b), and MIS 5e (Langford et al., Reference Langford, Boreham, Briant, Coope, Horne, Penkman, Schreve, Whitehouse and Whittaker2017). Candy et al. (Reference Candy, Schreve, Sherriff and Tye2014) compiled, compared, and discussed published palaeotemperature evidence based on single-proxy (beetle and ostracod) range reconstruction methods as well as vertebrate and plant indicator taxa from five British terrestrial sites assigned to MIS 11, and similarly compared evidence from British MIS 11, 9, 7, and 5e interglacial sites, but did not attempt any systematic refinement of the reconstructions. Candy et al. (Reference Candy, White and Elias2016) used reconstructions obtained from beetle- and ostracod-based methods, together with threshold temperatures of vertebrate and plant macrofossils, to determine “most probable” winter and summer palaeotemperature ranges for British Ipswichian/Eemian interglacial period (MIS 5e) sites.
Here we present the first multiproxy quantitative palaeotemperature range reconstructions for MIS 11 in Britain, using new results from Hoxne that combine beetle-, ostracod-, and chironomid-based methods with threshold temperatures of plant indicator taxa and additional evidence from molluscs and small vertebrates. In addition, we present and interpret oxygen and carbon isotope analyses of ostracod shell carbonate from Hoxne Stratum C. We pay critical attention to the likelihood that some components of Stratum C assemblages are reworked from underlying interglacial deposits. We explore the challenges and potential of multiproxy palaeotemperature reconstructions with the aim of establishing and demonstrating a robust, reproducible approach.
METHODS
A summary column with the main lithostratigraphic units sampled and a map showing the locations of sample areas and boreholes are shown in Figure 1; stratigraphic locations of samples are shown in Figure 2.

Figure 2. Stratigraphic context of samples from Hoxne referred to in the text and the Supplementary Material; see Fig. 1 for geographic locations.
Comparisons between different proxy methods facilitate the checking, validation, and refinement of palaeotemperature reconstructions. We combined three proxy methods based on invertebrate fossils: the chironomid transfer function (CTF) method (e.g., Brooks, Reference Brooks2006), the mutual ostracod temperature range (MOTR) method (Horne, Reference Horne2007), and the beetle mutual climatic range (BMCR) method (Atkinson et al., Reference Atkinson, Briffa, Coope, Joachim, Perzy and Berglund1986). In addition, we took account of plant indicator taxa and considered the qualitative implications of molluscs and small vertebrates. Details of each method are given below; sample-processing methods are described in the Supplementary Material, Section 1.
For the CTF method, a weighted averaging–partial least squares two-component, mean July air temperature inference model (Birks, Reference Birks, Maddy and Brew1995) was used to reconstruct the mean July air temperature from the fossil chironomid assemblage. This model has a root-mean-square error of prediction of 1.01°C and a coefficient of variation (r 2) of 0.91 and is based on a modern Norwegian calibration set of 154 lakes covering a mean July air temperature range of 3.5–16.0°C (Brooks and Birks, Reference Brooks and Birks2000, Reference Brooks and Birks2001, Reference Brooks and Birks2004; Self et al., Reference Self, Brooks, Birks, Nazarova, Princhu, Odland, Yang and Jones2011). Following Larocque (Reference Larocque2001), Heiri and Lotter (Reference Heiri and Lotter2001), and Quinlan and Smol (Reference Quinlan and Smol2001), only samples including more than 50 chironomid larval head capsules were used for palaeotemperature reconstruction. Results are presented to the nearest 0.1°C.
The MOTR method was used to reconstruct mean January and July air temperatures to the nearest 1.0°C. Species calibrations are mainly from Horne et al. (Reference Horne, Curry, Mesquita-Joanes, Horne, Holmes, Rodriguez-Lazaro and Viehberg2012a), based on the NODE (Non-marine Ostracod Distribution in Europe) database and additional records from OMEGA (Ostracod Metadatabase of Environmental and Geographical Attributes; Horne et al., Reference Horne, Holmes, Rodriguez-Lazaro, Viehberg, Horne, Holmes, Rodriguez-Lazaro and Viehberg2012b).
For the BMCR application we used previously published species lists: Coope (Reference Coope, Singer, Gladfelter and Wymer1993) gave detailed descriptions, interpretations, and palaeotemperature reconstructions of beetle assemblages from Stratum F (Anglian late glacial) and Stratum D (Hoxnian late temperate), while Stratum C results were first published (partially) by Ashton et al. (Reference Ashton, Lewis, Parfitt, Penkman and Coope2008) and are updated here with the addition of more detail. We provide revised palaeotemperatures for Strata F and D based on the overall species lists, while Stratum C results are provided sample by sample. Reconstructions of mean January and July air temperatures to the nearest 1.0°C were calculated with the BMCR method (Atkinson et al., Reference Atkinson, Briffa and Coope1987), using the BugsMCR function of the BugsCEP database (Buckland and Buckland, Reference Buckland and Buckland2012) to calculate the range of T max (July temperature) and T min (January temperature) for each sample/site investigated; T range determines the level of continentality. The technique has undergone several revisions in recent years (Buckland, Reference Buckland2007), providing updated results, while the sample-by-sample analysis for Stratum C provides significant new palaeoclimatic insights.
Pollen and plant macrofossil records of species indicative of threshold temperatures were obtained from West (Reference West1956) and are given to the nearest 0.5°C.
The taphonomic implications and palaeoclimatic significance of nonmarine molluscan and small vertebrate fossils are considered qualitatively.
We applied a multiproxy consensus (MPC) approach using overlapping ranges produced by different proxy methods to determine the narrowest range that agrees with all of the proxy results for that stratum, thus arriving at a “mutual mutual” temperature range (Langford et al., Reference Langford, Boreham, Briant, Coope, Horne, Schreve, Whittaker and Whitehouse2014a, Reference Langford, Boreham, Coope, Fletcher, Horne, Keen, Mighall, Penkman, Schreve and Whittaker2014b) here termed the “mutual overlap range” (MOR). Unlike the BMCR and MOTR methods, which reconstruct ranges (the actual temperature is considered to have been “somewhere within” that range), the CTF method produces a single “most probable” value with ± error bars (representing 1 SD each way), which for multiproxy comparisons are considered to represent a range. Results of the MPC are presented to the nearest 1.0°C.
In some cases in which not all proxy results initially overlapped, improvements were achieved by reconsideration of the assumptions and data on which the reconstructions are based. For example, revision of BMCR results using new calibrations has removed or reduced gaps that were apparent in our first attempts at multiproxy reconstruction. Where a gap remains, a “best fit” is obtained by establishing the narrowest temperature range that is in contact with all the proxy results for that stratum, even if this leaves gaps that are not covered by reconstructed values; this is termed a “minimum gap range” (MGR). Clearly, MGRs are much less certain than MORs, because the former, lacking complete sets of mutual overlaps, suggest errors or uncertainties in the method or the data. Such discrepancies highlight limitations of the multiproxy approach but at the same time provide valuable opportunities to reconsider some of the assumptions that underpin the methods. Indeed, we regard the recognition of such limitations as a strength rather than a weakness of the MPC approach, because it provides a measure of the reliability of the results.
Stable isotope analyses were carried out on specimens of Ilyocypris (“bradyi”-type) and Cytherissa lacustris from Stratum C; further details are given in the Supplementary Material, Section 1. The results are expressed in standard δ units on the VPDB scale with a mean standard deviation of measurements of 0.05‰ and 0.03‰ for δ18O and δ13C, respectively.
A list of full taxonomic names (genus, species, author, date) is provided in the Supplementary Material, Section 7.
RESULTS
Chironomid assemblages
Productive chironomid samples were obtained from Strata C and E (Figs. 1 and 2); results from those sufficiently abundant to justify further analysis are shown in Figure 3. Stratum E chironomid assemblages are relatively species poor, comprising taxa typical of warm, eutrophic waters with submerged aquatic vegetation; low oxygen conditions existed, either in a deep, thermally stratified lake or in a shallow lake containing large amounts of decaying vegetation. Stratum C chironomid assemblages are relatively species rich, indicative of relatively shallow, well-oxygenated waters with submerged aquatic vegetation, possibly influenced by stream inflow, and dominated by widespread, eurytopic taxa that can occur in both warm and temperate waters; they include cold stenothermic taxa, but the overall indications are of cool-temperate rather than cool climate. Further details are given in the Supplementary Material, Section 2.

Figure 3. Summary of chironomid occurrences recorded from MIS 11 strata at Hoxne.
It is possible that thermophilic taxa present in Stratum C (“Arctic Bed”) were reworked from earlier sediments deposited during a warmer period (e.g., Stratum E). Indeed West (Reference West1956, p. 313) suggested this possibility when considering the vegetation record at Hoxne, identifying the pollen spectrum of a clay mud pebble from Stratum C as being from HoIId (uppermost Early Temperate, within Stratum E; redesignated HoIIc by Turner and West [Reference Turner and West1968]). However, the Stratum E chironomid assemblages, which represent a warm climate, are dominated by Chironomus plumosus-type, and yet this taxon is hardly present in any of the Stratum C samples, which would be expected if there had been reworking of sediments. Equally, the temperate taxa that are well represented in the Stratum C sediments, especially Paratanytarsus penicillatus-type and Microtendipes pedellus-type, are absent from the samples analysed from Stratum E, suggesting these taxa have not been reworked.
CTF method palaeotemperature reconstructions
All reconstructed ranges are mean monthly air palaeotemperatures; the CTF method was used to reconstruct only July temperatures. Results are detailed in Table 2 and illustrated in Figures 3 and 4. Stratum E results are +19.1°C ± 1.4 and +19.3°C ± 1.8; equivalent to a combined range (MOR) of +17.8°C to +20.5°C. Stratum C results range from +13.4°C ± 1.1 to +15.0°C ± 1.1, equivalent to a combined MOR of +13.9°C to +14.5°C.
Table 2. Chironomid transfer function palaeotemperature reconstructions.a


Figure 4. Multiproxy palaeotemperature reconstructions for MIS 11 at Hoxne.
Ostracod assemblages
Ostracod assemblages were recovered from Strata F, E, C, and B (Figs. 1 and 2); details are given in the Supplementary Material, Section 3. The assemblages are composed entirely of typically freshwater taxa (Table 3), although many of those can tolerate slightly brackish waters, and are consistent with lacustrine and (in the case of Stratum B) fluvial depositional environments (Supplementary Material, Section 3).
Table 3. Summary of ostracod occurrences recorded from MIS 11 strata at Hoxne, with calibrations used in the mutual ostracod temperature range method where available.

a An asterisk (*) denotes extinct species.
MOTR method palaeotemperature reconstructions
All reconstructed ranges are mean monthly air palaeotemperatures. The calibrations used (Table 3) are from Horne et al. (Reference Horne, Curry, Mesquita-Joanes, Horne, Holmes, Rodriguez-Lazaro and Viehberg2012a), with two exceptions. In the case of C. lacustris, an adjustment was made to take account of living records in Lake Biwa, Japan (Smith and Janz, Reference Smith and Janz2009), raising the maxima of its mean January and July ranges by 1°C and 3°C, respectively. In the case of Limnocythere suessenbornensis, found in Stratum F and considered to be extinct in Europe, an unpublished calibration of the living North American species Limnocythere friabilis was applied, based on recent work by DJH with Alison Smith (Kent State University, Ohio, USA) that suggests that the two species are synonymous. Extinct species (Cyclocypris obunca, Ilyocypris quinculminata, Leucocythere batesi, Limnocythere falcata, Scottia browniana) are not included in the analyses, because they cannot be calibrated. Two living species are not used due to taxonomic uncertainties; attempted calibrations of Ilyocypris gibba are considered unreliable, because it has frequently been confused with other congeneric species, and there are insufficient living records (only two) of Paralimnocythere compressa in NODE and OMEGA to permit its calibration. At least three other species are present in our Hoxne assemblages, lumped together as Ilyocypris spp. in Table 3; one of these may be Ilyocypris bradyi Sars, 1890, another Ilyocypris decipiens Masi, 1905, while the third may be a new species, and further study will be necessary before their taxonomy and occurrences are well enough understood to include them in our analyses. All the reconstructions produced 100% overlap of calibrated ranges (i.e., no outliers with nonmutual ranges). Results are illustrated in Figures 4 and 5 and detailed in Table 4.
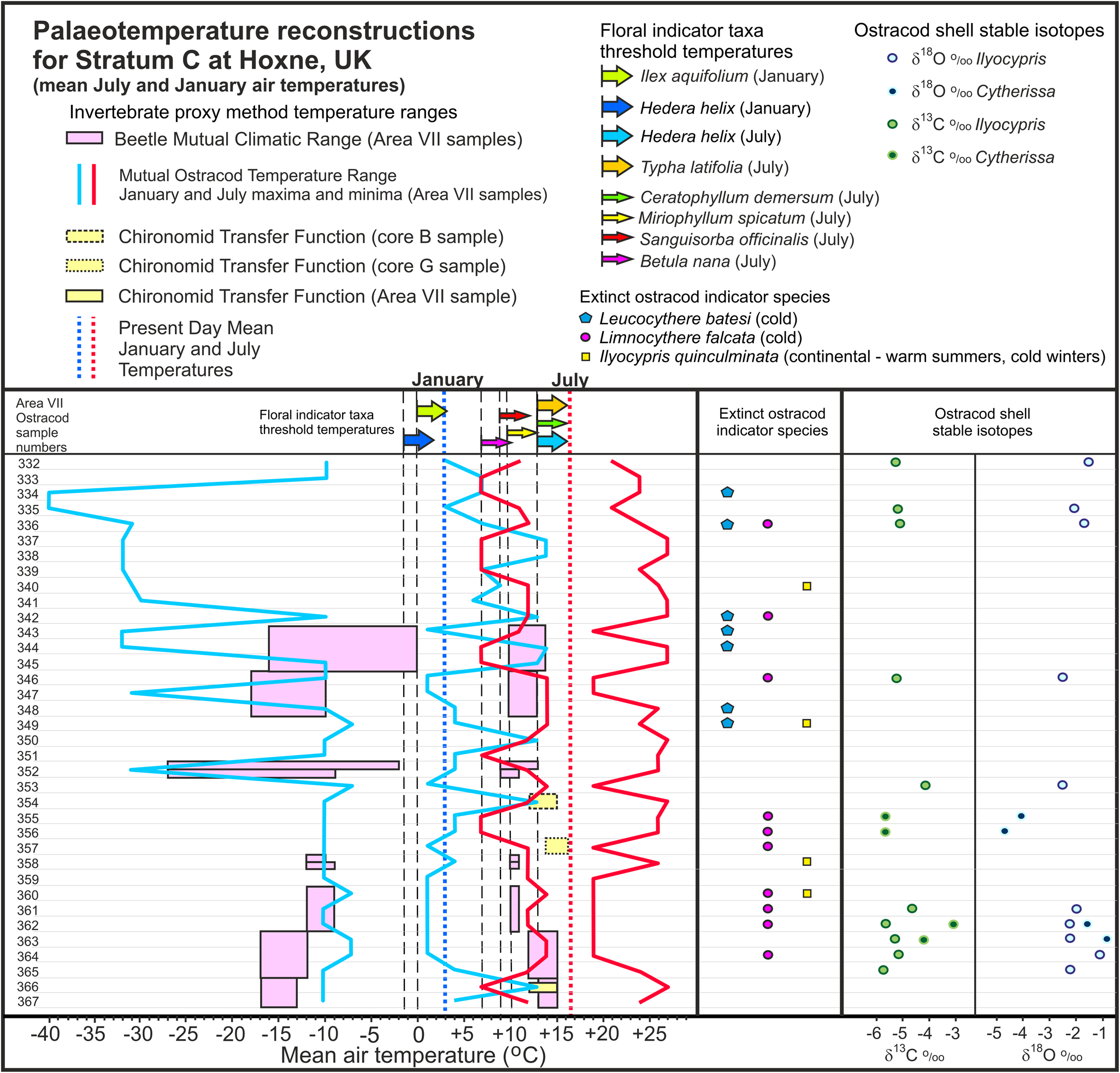
Figure 5. Stratum C, Hoxne: palaeotemperature reconstructions from mutual ostracod temperature range (MOTR), beetle mutual climatic range (BMCR), and chironomid transfer function (CTF) methods, with the addition of plant threshold temperatures and ostracod shell δ18O and δ13C measured in shells of Cytherissa and Ilyocypris. Note that the BMCR and CTF sample intervals were determined from a different numbering sequence matched to the equivalent ostracod sample numbers shown on the left of the figure (see Fig. 2 for details).
Table 4. Mutual ostracod temperature range results from Hoxne.a

Stratum F results for each of three samples are provided in Table 4; the MOTR ranges for the combined assemblage being January −8°C to −3°C and July +16°C to +23°C. Stratum E results from three separate sample assemblages are likewise given in Table 4, the combined MOTR reconstruction being January −8°C to +3°C and July +12°C to +21°C.
High-resolution sampling of Stratum C yielded ostracod assemblages from 36 consecutive samples, comprising a mixture of taxa commonly regarded as cold climate indicators (e.g., L. falcata, Limnocythere sanctipatricii, C. lacustris) and warm/temperate indicators (e.g., Metacypris cordata, Darwinula stevensoni). It cannot be ruled out that at least some of the warm-climate taxa were introduced into Stratum C by reworking from older (interglacial) lake sediments exposed around the margins during times of lowered lake level. However, because all the taxa included in the MOTR analysis do have overlapping, mutual ranges (i.e., they could have coexisted) and there are no reliable criteria (e.g., preservational differences) for distinguishing between in situ and reworked specimens, it had to be assumed that they are all in situ and thus valid for the MOTR reconstruction; potential problems of reworking are considered further in the “Discussion” section. Accordingly, MOTR reconstructions for Stratum C, based on 10 calibrated taxa, are presented in Figure 5 and Table 4. The sample-by-sample reconstructions appear to demonstrate considerable variation in climatic conditions through the sequence. Because the actual temperatures could be anywhere within the ranges reconstructed, it is possible that they could have stayed between narrower limits throughout the interval, with little or no variation, but comparison with the other proxy results suggests that this is unlikely.
The MOTR ranges for the combined Stratum C assemblage are January −7°C to +1°C and July +14°C to +19°C. The lower extreme of the July MOTR is 1° cooler than the lower extreme of Benardout's (Reference Benardout2015) July result for Stratum C (+15°C to +19°C), a discrepancy that can be explained by our decision not to include I. decipiens in our analyses because of taxonomic uncertainties within the genus Ilyocypris. Also worthy of note are the occurrences of three extinct taxa with palaeoclimatic significance: L. batesi, L. falcata, and I. quinculminata. The first two are considered to be cold-climate indicators by association with other taxa (Whittaker and Horne, Reference Whittaker, Horne, Whittaker and Hart2009). Ilyocypris quinculminata was considered to be a warm indicator by association with other taxa (Whittaker and Horne, Reference Whittaker, Horne, Whittaker and Hart2009), so its sporadic occurrence in Stratum C (and absence from the other strata) seemed anomalous. Because it is extinct, its temperature ranges cannot be calibrated, but two occurrences for which MOTR results have been published more recently offer an insight into its palaeotemperature tolerances. At Boxgrove (MIS 13), it was found in an assemblage that provided palaeotemperature reconstructions, based on a combination of MOTR and herpetile-mutual climatic range (MCR) methods, of −4°C to +4°C (January) and +15°C to +20°C (July), which in comparison with modern temperatures for that vicinity are consistent with similar summers but suggest colder winters (by at least 2°C) than today (Holmes et al., Reference Holmes, Atkinson, Darbyshire, Horne, Joordens, Roberts, Sinka and Whittaker2010). In MIS 11 at Ebbsfleet (Southfleet Road), it occurred in association with extant ostracod taxa that together yielded MOTR values of −4°C to −1°C (January) and +17°C to +21°C (July) (Whittaker et al., Reference Whittaker, Horne, Wenban-Smith and Wenban-Smith2013), which for that region suggests warmer summers but colder winters (by at least 4°C) than today (a more continental climate). We conclude, tentatively, that I. quinculminata may be regarded as an indicator of a temperate/cool continental climate similar to that of northern Germany and Poland today, with (in comparison with the maritime/oceanic climate of present-day England) warmer summers and colder winters.
Stratum B1 ostracods were sparse, yielding wide-ranging MOTR results from two samples (Table 4); the combined MOTR reconstructions are January −8°C to +13°C and July +10°C to +25°C.
Beetle assemblages
Productive beetle samples were obtained from Strata C, D, E, and F (Figs. 1 and 2). The beetle assemblage of Stratum F was obtained and described by Coope (Reference Coope, Singer, Gladfelter and Wymer1993) from a single sample of laminated drift mud collected at the base of the sequence in West's (Reference West1956) section 100 (Fig. 1). Stratum F yielded 82 taxa, of which 69 could be named to species, the assemblage being dominated by cold-adapted species (e.g., Pycnoglypta lurida, Holoboreaphilus nordenskioeldi, Tachinus caelatus, Simplocaria metallica). There were also more temperate, less cold-adapted species present, such as Bembidion guttula and Colymbetes fuscus, although the ranges of both of these extend north of the Arctic Circle in Fennoscandia at the present day.
Stratum E beetle samples from BH 02/1 (Figs. 1 and 2) yielded only sparse assemblages, likely due to small sample size. The Stratum D beetle assemblage, originally published by Coope (Reference Coope, Singer, Gladfelter and Wymer1993), is from a single block of woody peat taken from Cutting XXIII of the 1971–1974 excavations (Wymer, Reference Wymer, Singer, Gladfelter and Wymer1993; Fig. 1); it is very diverse and clearly associated with temperate conditions.
Stratum C beetle assemblages were obtained from Area VII samples (Figs. 1 and 2) and first published by Ashton et al. (Reference Ashton, Lewis, Parfitt, Penkman and Coope2008). Table 5 shows critical species from Stratum C that have significance because none of them lives in Britain today. Two climatically significant species in this assemblage are not represented in the BugsCEP database; Hippodamia arctica is a high boreal species living in Fennoscandia and northern Russia with an outpost in the Caucasus Mountains (Brundin, Reference Brundin1934), where it is found on Betula nana–rich Empetrum heath, while S. metallica is a boreo-alpine species, ranging from Greenland and Fennoscandia and the high mountains of central Europe (Horion, Reference Horion1953), that feeds exclusively on mosses.
Table 5. Calibrated present-day mean monthly temperature ranges of selected beetle taxa from Stratum C, Hoxne.a

a Data based on BugsCEP envelopes with the exception of P. lurida and O. boreale, which are based on updated envelopes generated using GBIF, which demonstrate improved winter temperature envelope values (Milne, Reference Milne2016). Only a restricted number of revised envelopes are available via this source.
It is interesting to observe the presence in Stratum C of a few species that also occur in the underlying Stratum D, suggesting the possibility of reworking. It is noteworthy that the lowermost assemblages of Stratum C (374 and 373; Table 6) are cool temperate, indicating warmer summer conditions than the rest of the sequence, but still very cold winter conditions; there is a small assemblage of wood/tree-associated species (3.6% of the terrestrial fauna) as well as a number of dung beetles and species associated with open ground (4.5% of the terrestrial fauna). The abundance of individuals and diversity of taxa in these samples is more typical of cool-temperate conditions, including a range of ground beetles, hydrophilid and staphylinid beetles, leaf beetles, and weevils. A wide and diverse range of aquatic and semi-aquatic vegetation is indicated by the reed beetles, not seen again higher in the sequence. These include Donacia dentata, which is found on Sagittaria sagittifolia and occasionally Potamogeton spp. (Hyman, Reference Hyman1992); it is not an alpine taxon, and its distribution in Britain today is largely southern (Hyman, Reference Hyman1992; Koch, Reference Koch1992); similarly, Plateumaris affinis is not an alpine species and, along with Donacia thalassina, is found on club rushes and sedges (Carex spp., Scirpus spp.) (Hyman, Reference Hyman1992; Koch, Reference Koch1992). Donacia semicuprea is found on Glyceria spp. (Bullock, Reference Bullock1993). Donacia aquatica cocoons on Ranunculus lingua, Sparganium simplex, Glyceria spp., and Carex spp. (Stainforth, Reference Stainforth1944), similar habitat for Donacia cinerea. This rich waterside vegetation fauna is also found in Stratum D, although in a more abundant and diverse assemblage. There are also, however, several species that indicate very cold winter conditions, including Boreaphilus henningianus and H. nordenskioeldi.
Table 6. Beetle mutual climate range results from Hoxne.a

The other species common to Strata C and D is the saproxylic Stenoscelis submuricatus, which appears in both lowermost samples from Stratum C. Today, it lives in the sapwood of Populus and Salix, but also on Alnus, Fraxinus, and Fagus (Koch, Reference Koch1992). It is clearly a temperate species that has been found in other Hoxnian sites (e.g., Nechells, Warwickshire) as well as MIS 7, 9, and 5e sites. It was very abundant at the top of Stratum D. Its presence in Stratum C may seem anomalous but is not necessarily inconsistent with the assemblage, as its ability to live on a variety of tree hosts (including Salix, notwithstanding that Salix polaris is a tundra shrub rather than a tree) may have allowed it to be more tolerant of cool conditions. The original view, based on research by Gaunt et al. (Reference Gaunt, Coope, Osborne and Franks1972) and Hoffmann (Reference Hoffmann1954), was that this is a Mediterranean species, but more recent records show that it can also be found in western and eastern Europe across to Switzerland, Austria (Folwaczny, Reference Folwaczny, Freude, Harde and Lohse1983), Hungary (Szénási, Reference Szénási2014), montane regions of France and the Pyrenees (Hoffmann, Reference Hoffmann1954), and northern coastal France (GBIF, 2018). A review by Whitehouse (Reference Whitehouse2006) of many non-British Holocene fossil cossinine weevils, a group to which S. submuricatus belongs (note that some authors prefer to assign the species to the genus Rhyncolus), suggests their modern distribution is more likely to be influenced by availability of quality habitat rather than temperature per se. Recent species distribution models of Rhyncolus and other saproxylic taxa (Whitehouse, N., Milne, G., Cameron, A., unpublished data) suggest that many appear responsive to continental conditions and are especially adversely impacted by wetter conditions rather than cold or warm temperatures per se (see further discussion on Rhyncolus, in the “BMCR Method Palaeotemperature Reconstructions” section). They are, however, always associated with tree habitats. Several other species attest to the presence of trees, including Agrilus sp., whose larvae develop under the bark or wood of deciduous trees, while Eledona agricola lives on various fungi growing on deciduous trees, principally Polyporus sulphuraeus on species of Salix. There are, therefore, grounds for reconsidering whether S. submuricatus signifies warm conditions; its biology and ecology could be more in keeping with the rest of the faunal assemblage, indicative of cooler summer temperate conditions and cold winter conditions. There are three species typical of northern boreo-montane conditions, including P. lurida, Helophorus obscurellus, and S. metallica (the first two not now living in the British Isles). Simplocaria metallica is found in montane and alpine areas of central Europe (Koch, Reference Koch1989); while P. lurida is found as far south as Denmark, Germany, and Poland (Koch, Reference Koch1989; Gusarov, Reference Gusarov1995); and H. obscurellus is an Asiatic species, found as far west as the Kanin Peninsula, Russia (Coope, Reference Coope1994). None is associated with high arctic conditions, and all have been found associated with late glacial deposits during cooler interstadial periods; P. lurida, for instance, can be found in summer warmth up to +24.5°C and winter temperatures as mild as −0.4°C (Table 5). They are not inconsistent with cooler temperate conditions indicated by the rest of the assemblage, suggesting continental conditions of cold winters but moderately warmer summers during the early stages of Stratum C, compared with later samples from this stratum.
In summary, despite some ecological similarities between Stratum C samples 374–373 and Stratum D, on balance, those from Stratum C appear to constitute a coherent in situ assemblage, rather than one contaminated with reworked material, indicating cold winter temperatures and cool summer temperatures of sufficient spring and summer warmth to allow some tree growth, perhaps in the form of Salix and other cool-tolerant species (e.g., Betula) in the basal portions.
Additional notes on selected climatically significant species from Strata F and C (none of which occurred in Strata E or D) are given in the Supplementary Material, Section 4.
BMCR method palaeotemperature reconstructions
The BMCR results are illustrated in Figures 4 and 5 and summarized in Table 6.
Previously published BMCR results for Stratum F, based on 24 species, were January −26°C to −9°C and July +10°C to +12°C (Coope, Reference Coope, Singer, Gladfelter and Wymer1993). To explain the mixture of taxa (warm and cold) from this interval, Coope (Reference Coope, Singer, Gladfelter and Wymer1993) discussed the possibility that some were reworked, but considered it likely that the mixture was due to fairly rapid climate change in the interval represented by Stratum F, with July temperatures initially about +10°C but quickly rising to +15°C. However, rerunning Coope's (Reference Coope, Singer, Gladfelter and Wymer1993) list in BugsCEP (36 species, 89% overlap, which means 11% had outlier palaeotemperature ranges that were not mutual with the 89%) produced revised reconstructions suggesting substantially warmer climate (January −11°C to −4°C, July +15 to +16°C). Failure to find 100% overlap likely supports the idea that this interval encompassed a period of rapid climate change, such that the sampled assemblage does not represent a single snapshot of stable climate.
Stratum E yielded only nine beetle species from four small samples of the lacustrine sediments, and because so few predatory or scavenging species were found, no MCR reconstruction could be made. However, they include several species, not in the MCR database, which indicate that the temperatures at the time were at least as warm and possibly warmer than those of the present day and likely to be reasonably continental. Notable amongst these is the non-British Rhyncolus elongatus, a saproxylic species associated with decaying, rotting pine (Koch, Reference Koch1992). Its distribution is today in central and eastern parts of northern and central Europe up to southern Fennoscandia (Whitehouse, Reference Whitehouse1997, Reference Whitehouse2000). Recent exploratory research on this species and other members of this genus by N. Whitehouse, G. Milne, and A. Cameron (unpublished data), using the Maxent software maximum-entropy approach for modeling species niches and distributions, with 18 bioclimatic variables, supports the idea that its predicted climatic niche based on modern distributions is largely in eastern Europe and southern Sweden. Temperature seasonality appears to be the strongest driver of its predicted climatic niche, followed by mean diurnal range and minimum temperatures of the coldest month. This suggests that while seasonality is an important determinant of the distribution of R. elongatus, it is also sensitive to daytime temperature extremes and winter cold. Another species in the assemblage supports temperate conditions: the deathwatch beetle Xestobium rufovillosum is not found in its natural environment, represented by present climate, in northern Britain (Alexander, Reference Alexander1994).
The originally published BMCR results for Stratum D (Coope, Reference Coope, Singer, Gladfelter and Wymer1993; Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008) were January −10°C to +6°C and July +15°C to +19°C. The revised results, based on 25 species, are January −10°C to + 1°C and July +16°C to +19°C (96% overlap), showing a July lower limit that is 1°C higher and a January upper limit 5°C lower than the original ones. The suboptimum overlap is likely due to minor in-sequence climate change (although there is no obvious evidence for this in the species list) and/or because the full climate range of one or more species has not been fully captured (Milne, Reference Milne2016).
Stratum C reconstructions using BugsCEP are presented on a sample-by-sample basis (Fig. 5, Table 6), as well as overall (Fig. 4), using the combined assemblages to represent Stratum C. Using 23 species of predatory or general scavenging beetles, combined BMCR results are January −16°C to −2°C and July +12°C to +14°C (82.6% overlap), indicative of somewhat warmer (though still markedly colder than present-day) summer conditions than quoted by Ashton et al. (Reference Ashton, Lewis, Parfitt, Penkman and Coope2008) (July at or <10°C) but consistent with their winter reconstruction (January about −15°C). The fact that 100% overlap was not achieved suggests that climate variation occurred during the times represented by the relatively coarse sampling interval, and this is borne out by the BMCR sample-by-sample reconstructions (Fig. 5) and supported by evidence from the high-resolution ostracod results described earlier.
The sample-by-sample reconstruction for Stratum C (Fig. 5, Table 6) demonstrates clear differences in climatic conditions through the sequence. Working up from the base of the sequence, where summer temperatures were only slightly cooler (+13°C to +15°C in sample 374) than today, we see cooling to +9°C to +11°C (sample 369), a mean July temperature drop of at least 2°C alongside a slight amelioration of winter temperatures through the same interval, with the upper limit of the January range rising from −13°C (sample 374) to −9°C (sample 369). Also noteworthy is the step between the reconstructions for samples 373 and 372, which show no overlap of their reconstructed temperature ranges, indicating a significant and relatively rapid change involving summer cooling and winter warming and suggesting a transition from continental to oceanic climate. It must also be noted, however, that the size of the winter envelopes is large for these samples and extends to very low values. The coldest envelopes are particularly controlled by two species that can tolerate extremely cold winters, Boreaphilus henningianus and H. nordenskioeldi, both of which are recorded as being associated with winter temperatures as low as −34°C and −35°C, respectively (Table 5). It is impossible to determine where exactly within an envelope the true temperature was situated, but it seems likely, given these species also occur in adjacent assemblages that give slightly warmer reconstructed temperatures, that conditions were never as cold as the lowest limits that the reconstructions might imply. The upper samples provide reconstructions that suggest climatic warming, with ranges of January −16°C to 0°C and July +10°C to +14°C indicated by the topmost assemblage (sample 328). Note that combining the sample-by-sample results to obtain overall representative ranges for Stratum C produces MGRs (January −13°C to −12°C, July +11°C to +13°C) that differ from those obtained by applying the BMCR method to the combined assemblage from all of the Stratum C samples (January −16°C to −2°C, July +12°C to +14°C), which produced an overlap of only 82.6%; this demonstrates that low sampling resolution may have merged short-term climatic changes to which the beetle taxa responded, resulting in gaps between the calibrated ranges of some included taxa and thence a lower overlap percentage for the whole assemblage.
Importantly, this BMCR reconstruction does not use plant-associated species and is therefore independent of (as well as consistent with) the overall beetle assemblage qualitative evidence (discussed earlier). The diversity of wood-loving taxa evident in Stratum D is absent from the rest of Stratum C, and conditions are cooler than Stratum D, becoming increasingly cooler over time (although S. submuricatus reappears in the upper portions of the sequence); therefore, the Stratum C assemblages represent an increasingly cooling community that is largely distinct from the temperate community represented by Stratum D.
In summary, the sample-by-sample reconstruction suggests a previously unknown summer cooling event coincident with winter amelioration within Stratum C. However, its registration within winter temperatures is much less clear due to the much larger envelope sizes generated by the present BugsCEP MCR package. New envelopes that have recently been developed and tested independently using modern modeling techniques (Milne, Reference Milne2016) suggest that the current envelopes tend to overestimate winter cold and summer warmth when compared against 12 modern test sites and species lists. The new envelopes are not yet fully operational, so are not used here.
Finally, the types of species used for each sample BMCR suggest the reconstructions are robust. A common assumption of the BMCR methodology has been that increasing the number of taxa used in a reconstruction will give a more precise (i.e., narrower) temperature range (Atkinson, T., personal communication, 2013). Modern testing work has shown that there is no discernible relationship between the number of taxa used in the BMCR reconstruction and the obtained BMCR temperature ranges for summer or winter (Milne, Reference Milne2016), but that understanding species niche traits is crucial. As Russell Coope highlighted both vocally and in print, “species should not be treated merely as ciphers in some purely mathematical model” (Coope, Reference Coope2010, p. 1511); thus, simply including more beetles in a reconstruction will not necessarily produce a better result. What may be more pertinent is whether the species are eurytherms (climate generalists) or stenotherms (climate specialists): the type of species used in a reconstruction has huge importance for the output. It is thus vitally important that users of any reconstruction system understand the climatic and ecological conditions associated with the taxa used in order to understand the robustness of a reconstruction. In this reconstruction, a high number of stenotherms have been used, increasing the confidence levels of the reconstruction.
Palaeoclimatic implications of flora
Several plant indicator taxa recorded at Hoxne (West, Reference West1956) are indicative of threshold temperatures (Figs. 4 and 5). Of the pollen taxa, Hedera (assumed to be Hedera helix; West, Reference West1956) requires mean air temperatures of −1.5°C or higher in January and +13°C or higher in July (Iversen, Reference Iversen1944; West, Reference West1956). Ilex (assumed to be Ilex aquifolium; West, Reference West1956) requires a mean January temperature of at least −0.5°C or 0°C (the latter value is used herein) (Iversen, Reference Iversen1944; West, Reference West1956; Walther et al., Reference Walther, Berger and Sykes2005). Typha latifolia needs a mean July temperature of +13°C or above (Isarin and Bohncke, Reference Isarin and Bohncke1999). All three taxa occur in Strata C, D, and E at Hoxne, with the last mentioned being found additionally in Stratum F (West, Reference West1956). Minimum mean July air temperature requirements of four plant macrofossil species were obtained from Isarin and Bohncke (Reference Isarin and Bohncke1999): B. nana (+7°C), Ceratophyllum demersum (+13°C), Miriophyllum spicatum (+10°C), and Sanguisorba officinalis (+9°C). All four occur in Stratum C, and C. demersum also occurs in Strata D, E, and F (West, Reference West1956).
Another potentially significant plant taxon, the water fern Azolla filiculoides, was commented on by West (Reference West1956), who found it commonly in Stratum C (but suggested that it may have been reworked from older strata), abundantly in Stratum D, and occasionally in the middle and upper parts of Stratum E. He considered its palaeoclimatic value doubtful, as it is widely distributed in the Americas and has an “erratic” behaviour in western Europe, and he noted that it survives under ice but is killed by winter frosts. On the other hand Hall (appendix A in Coxon, Reference Coxon1985) noted the occurrence of Azolla megaspores at nearby Athelington as being of particular interest, as it is typical of Hoxnian zones II and III, and because he considered it indicative of somewhat higher summer temperatures than at the present day. Experimental evidence has shown that adult plants survived at −2°C but were killed at −4°C in the laboratory, but they can survive air temperatures (or being encased in ice) of −5°C or even lower in the wild, given the right microclimatic conditions (Janes, Reference Janes1998). It apparently grows best at +15°C to +20°C and high irradiance (Janes, Reference Janes1998), although its optimum temperature for nitrogen fixation and oxygen evolution is +25°C (Wong et al., Reference Wong Fong Sang, Van Vu, Kijne, Thanh Tam and Planque1987). Populations in Poland have been observed to survive frosts down to −22°C (Szczęśniak et al., Reference Szczęśniak, Błachuta, Krukowski and Picińska-Fałtynowicz2009). In the light of this evidence, its status as a thermophile in the context of the British Quaternary is questionable, to say the least; consequently, we have not included it in our analyses.
Molluscan assemblages
Nonmarine molluscs were obtained from the same samples processed for other proxies as part of the Ancient Human Occupation of Britain (AHOB) project excavations (Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008), from Strata E, D, C, and B2 (Figs. 1 and 2); see the Supplementary Material, Section 1, for further details. The first analysis of nonmarine molluscan remains spanning the complete Hoxne sequence was undertaken by White (Reference White2012), and relevant data are published here for the first time (Table 7). Stratum F data from the quantified faunal list of Sparks (Reference Sparks1956) have been included to complete the Hoxne molluscan succession, but it should be noted that they came from a more marginal part of the lake than the other material.
Table 7a. Freshwater Mollusca from Hoxne, Suffolk; relative abundance (%) values used to plot Fig. 6.a Note that Table 7 is in four parts: 7a–d.




aData from Sparks (1956) and White (2012). See Figs. 1 and 2 for locations and stratigraphic context of samples.
The Stratum F freshwater assemblage is dominated by gastropods, namely Gyraulus laevis (35.8%) and Gyraulus crista (34.4%), with subordinate numbers of Valvata piscinalis (8.0%) and Radix balthica (7.8%). Hippeutis complanatus is also well represented (3.9%). Several other gastropod taxa were recorded at low frequencies. Bivalves, predominantly Pisidium nitidum, account for only 9.8% of the overall assemblage. The molluscan evidence from Stratum F is indicative of a relatively shallow lacustrine environment, rich in aquatic vegetation and inhabited by species tolerant of colder conditions (White, Reference White2012).
Assemblages from the overlying sediments of Stratum E are dominated by the freshwater gastropod V. piscinalis, which constitutes more than 90% of most samples and is indicative of a relatively deep-water habitat. Also present at low frequencies throughout Stratum E is the extinct bivalve Pisidium clessini, which occurs in several middle Pleistocene interglacial periods and is a member of the “Rhenish suite” of aquatic molluscs characteristic of Hoxnian pollen substage HoIIb (White et al., Reference White, Preece and Whittaker2013). None of the molluscan taxa present within Stratum E are obligate thermophiles, but all are compatible with the temperate climate indicated by the Hoxne pollen record (West, Reference West1956; Turner, Reference Turner1970).
The molluscan assemblage of Stratum D includes V. piscinalis, Bithynia tentaculata, Anisus leucostoma, and indeterminate species of Pisidium (Fig. 6). The shells from this part of the sequence were in relatively poor condition, with a distinctive dark stain indicative of the peaty sediments from which they were recovered. It is not clear whether this assemblage represents an in situ fauna or material derived from the underlying lacustrine deposits. Only the early part of the period represented by Stratum D, during which lake water levels were still reasonably high (cf. Coope, Reference Coope, Singer, Gladfelter and Wymer1993), would have provided a suitable habitat for V. piscinalis and B. tentaculata.

Figure 6. Selected freshwater Mollusca from Hoxne Strata F to B.
Mollusc shells recovered from Stratum C in Area VII were relatively poorly preserved, and a large proportion could not be identified to species level, leading to underrepresentation of species. The assemblages from Stratum C contain no obligate cold-adapted species but can be easily distinguished from the underlying temperate faunas from the earlier parts of the Hoxne sequence by the dominance of various species of Pisidium and the reduced abundance of V. piscinalis (Fig. 6). Similar bivalve-dominated faunas have been reported from other British cold-stage localities, such as Beetley (Preece, in West, Reference West1991), and in modern situations such as Scandinavian lakes (Kuiper et al., Reference Kuiper, Økland, Knudsen, Koli, Proschwitz and Valovirta1989; Økland, Reference Økland1990). Shells representing species common in the underlying sediments, such as Valvata cristata and B. tentaculata, were probably reworked from older deposits, although alternative interpretations are possible (see “Discussion: Reworking in Stratum C Assemblages”). However, several bivalve species present in Stratum C, notably Pisidium amnicum and Pisidium henslowanum, can be regarded as part of an in situ fauna, because with the exception of single specimens of the latter recorded in Stratum F and the basal sample of Stratum E, they are absent from the earlier part of the Hoxne sequence (Fig. 6).
In the fluvial deposits of Stratum B2, the molluscan assemblages include a more diverse gastropod fauna dominated by V. piscinalis and B. tentaculata and are broadly similar to the temperate fauna encountered in Stratum E (Fig. 6).
Calculation of the ratio of Bithynia shells to opercula has proved to be a useful means of assessing the energy dynamics of depositional environments (e.g., potentially discriminating between lacustrine and fluvial conditions) and can contribute to assessment of reworking. At Hoxne, the ratio of Bithynia shells to opercula within Strata F and E is balanced at close to 1:1 in most samples, indicative of minimal postmortem sorting and suggestive of a low-energy depositional environment consistent with a lake. In contrast, ratios of 1:3 and 1:4 typify samples recovered from Stratum C and may be considered to support the idea that Bithynia is a reworked element in these assemblages. Stratum C Samples in which only opercula are represented might indicate some decalcification of the sediments. It is also possible, however, that the Bithynia in Stratum C are in situ but were subject to postmortem sorting in the lake environment. Alexandrowicz (Reference Alexandrowicz1999) described a process whereby shells of dead Bithynia float for a more prolonged period of time than the opercula, which detach more quickly and sink, while the shells are driven shoreward by wind and wave processes. In the fluvial Stratum B2, evidence for higher-energy conditions is provided by Bithynia shell:opercula ratios of 1:10 (White, Reference White2012). These interpretations should be treated cautiously, as the numbers on which the shell:opercula ratios are based are generally very low (20 specimens or fewer) in samples from Strata E, D, and C, although many of the assemblages from Stratum B2 are big enough (>50) for the ratios to be considered more reliable.
Climatic significance of the molluscan assemblages
The molluscan record from Hoxne does not provide strong evidence for significant climatic variation due to the relatively broad tolerances of most of the taxa represented. Moreover, as noted earlier, several features of the record provide critical evidence relating to the potential for reworking and the taphonomic integrity of assemblages from Stratum C.
The most climatically significant taxon present in Stratum F is the gastropod G. laevis, which is characteristic of late-glacial or early postglacial molluscan faunas. It is common in the “Arctic Fresh-water Bed” at Ostend, Norfolk (Parfitt et al., Reference Parfitt, Coope, Field, Peglar, Preece and Whittaker2010), and a fauna dominated by G. laevis was reported in association with an early postglacial pollen profile at Tottenhill (Ventris, Reference Ventris1996). Similar freshwater assemblages dominated by G. laevis and G. crista have been documented from both late Anglian/early Hoxnian and late Devensian/early Holocene sequences (Kerney, Reference Kerney and Shotton1977). Given that the molluscan evidence from Stratum F is derived from a single bulk sample, it is difficult to compare with the data provided by the beetle faunas from this part of the Hoxne sequence, but they are not incompatible.
In the overlying temperate lacustrine deposits of Stratum E there are no obligate cold- or warm-adapted species that would place significant constraints on palaeoclimatic reconstructions. One species that provides some evidence for prevailing temperatures is V. cristata, which is not known from any Pleistocene cold-stage deposits. However, it occurs only as a rare element of the molluscan assemblages from Stratum E due to the deep-water environment represented by this unit, which is suboptimal for V. cristata. The limited molluscan fauna preserved in the Stratum D sediments provides little in the way of climatic information.
The molluscan assemblages from Stratum C contain no obligate cold-adapted species but can easily be distinguished from the preceding parts of the Hoxne sequence by the dominance of various species of the bivalve genus Pisidium. Similar faunas have been reported from other cold-stage localities such as Beetley (Preece, in West, Reference West1991) and in modern habitats such as Scandinavian lakes (Kuiper et al., Reference Kuiper, Økland, Knudsen, Koli, Proschwitz and Valovirta1989; Økland, Reference Økland1990). The presence of V. cristata in Stratum C at higher frequencies than the underlying Stratum E deposits has been attributed to reworking (cf. White, Reference White2012), although the rarity of this species in the upper part of the Stratum E deposits suggests that these shells might have been derived from sediments now missing from the sequence (i.e., the hiatus between Strata D and C). Alternatively, if they are genuinely part of the Stratum C fauna, their occurrence together with cold-tolerant taxa requires explanation. Another gastropod species known only from interglacial and interstadial sequences in Britain is B. tentaculata; this taxon is primarily represented in Stratum C by opercula, suggestive of reworking, although as noted earlier, other explanations are possible.
The assemblages from Stratum B2 are very similar to those from Stratum E, indicating a return to temperate climatic conditions. The similarity of the assemblages in these units is somewhat surprising, given the marked sedimentologic differences indicative of lacustrine (Stratum E) and fluvial environments (Stratum B2). The absence of P. henslowanum from Stratum B is noteworthy, given than it is a common species within Stratum C and prefers moving-water environments.
Small vertebrate assemblages from Stratum C
Small vertebrates were obtained from successive samples through Stratum C (Figs. 1 and 2); additional specimens were extracted from larger subsamples collected for insect remains (see the Supplementary Material, Section 1, for further details). A summary of the small vertebrates and other fossils recovered from the sieved samples is given in Table 8 and Figure 7. Bone fragments are not distributed uniformly through the sequence, with concentrations of bones in some horizons separated by samples that are virtually devoid of vertebrate material. Fish remains are the most abundant component, with identified specimens that include scales of Perca fluviatilis (perch), a tooth of Esox lucius (pike), pharyngeal teeth of cyprinids (carp family), and the distinctive spines of Pungitius pungitius (nine-spined stickleback). The bulk samples processed for insects yielded tiny pharyngeal bones of Alburnus alburnus (bleak) and Phoxinus phoxinus (minnow). Taken together, the general composition, low diversity of identified species, and relatively small size of the fishes imply a small waterbody, such as a pool or backwater.

Figure 7. Abundance of small vertebrate and other fossil remains in Stratum C (Area VII). Conspicuous organic seams correspond to peaks in the abundance of plant and other fossils remains in samples 314, 311, 308–307, and 304. Chalk stringers and lenses of chalky and gravelly sand occurring toward the top of the sequence (equivalent to samples 304–301) are associated with reworked Chalk Group fossils (corals, ostracods), pre-Quaternary fish remains, and marine molluscs. Occasional Mesozoic ostracods and foraminifera are also found in the lower organic-rich horizons.
Table 8. Small vertebrate remains from Stratum C, Area VII sample column, Hoxne.a

Only a few small mammal specimens were recovered from Stratum C. Among the more complete specimens, the presence of Dicrostonyx sp. (collared lemming) was determined from a molar fragment (sample 304) and a second upper molar (M2) fragment (sample 301). At least one other microtine rodent (vole) is represented by material too fragmentary to identify more precisely. For further details and discussion see the Supplementary Material, Section 5.
Taphonomy and environmental implications of the Stratum C vertebrates
Taking the assemblage as a whole, the vertebrate species recovered from Stratum C appear to be climatically coherent. The assemblage includes Dicrostonyx, a cold-indicating lemming of tundra and mammoth steppe (Guthrie, Reference Guthrie1968), together with fish species (P. fluviatilis, E. lucius, P. pungitius) that have geographic ranges that today extend above the Arctic Circle. The cyprinids could also belong to the same cold-stage assemblage. Although many cyprinid species are restricted in their ability to colonise the far north due to insufficiently high summer water temperatures in order to spawn successfully (Wheeler, Reference Wheeler1977), other cyprinids (including bleak, A. alburnus, and minnow, P. phoxinus, both found in Stratum C) can survive for long periods in ice-covered water and are found in rivers and lakes in the tundra, some as far north as the Arctic Ocean. The closest region to Hoxne where the geographic ranges of these taxa overlap today is located at the southern limit of the tundra south of the Kanin Peninsula. Today, this area is characterised by extremely cold winters (mean air temperatures of the coldest month between −14°C and −8°C) and summers with temperatures of the warmest month between +7°C and +14°C. Whether these estimates can be applied to Stratum C, however, depends on two critical assumptions: first, that the vertebrate assemblage represents animals that lived in the vicinity of the site during the period of accumulation of Stratum C, and second, that conditions did not change significantly during the deposition of this unit.
The first of these assumptions was investigated by examining the stratigraphic occurrence of different elements of the fossil assemblages recovered from the bulk samples (Fig. 7). Sedimentologically, Stratum C consists of silty muds interspersed with thin layers of organic detritus and, in its upper part, lenses of chalky gravel and sand. These coarser horizons represent episodic pulses of erosion and redeposition of older sediments exposed on the margins of the basin. These sediments contain a mix of fossil material, some of which (Mesozoic corals, ostracods and foraminifera, and Cenozoic marine molluscs) have undergone multiple cycles of transportation and redeposition, with initial erosion from the bedrock, transportation by glacial processes, and winnowing before burial at the depositional site. Plio-Pleistocene “crag” and glacial deposits (Lowestoft Till and “Corton beds”) at the edge of the Hoxne basin (Lord, Reference Lord and Dixon2012) provide a probable source for these reworked fossils. The horizons in Stratum C containing reworked pre-Quaternary fossils also correspond to peaks in the abundance of Quaternary fish, small mammals, and plant macrofossils, the last including well-preserved seeds and wood fragments up to 5 cm in diameter. This mix of debris appears to have been transported by pulses of higher-energy currents that washed the detritus into the basin. Marginal exposures of the Hoxnian lake muds provide a possible source for the Quaternary plant macrofossils (Stratum D) and fish remains (Stratum E). However, there is no obvious difference in the state of preservation (color differences, weathering, abrasion) of the vertebrate remains that identifies a reworked component in the Quaternary assemblage. With respect to taphonomy, while a few fish specimens in the Stratum C assemblage may be reworked from the interglacial deposits, the dominance of taxa (pike, perch, nine-spined stickleback) that are otherwise rare in the earlier lake muds suggests that the Stratum C accumulation reflects an essentially indigenous fauna. Similarly, the lemming teeth are unlikely to have been reworked from the interglacial sediments, and their presence in Stratum C plausibly indicates severe climatic conditions, at least during the deposition of the upper part of that unit.
Stable isotopes
Details of δ18O and δ13C measurements of ostracod (Ilyocypris and Cytherissa) shells from Stratum C are shown in Figure 5 and Table 9. The oxygen isotope composition of lacustrine carbonate is controlled by the temperature and isotopic composition of lake water. The isotopic composition of lake water, in turn, is determined by the composition of rainwater and inputs from groundwater or surface runoff, catchment effects (sensu Gat and Lister Reference Gat, Lister, Frengel, Stauffer and Weiss1995), and any evaporative enrichment. For ostracod shells, there is generally a positive offset from oxygen isotope equilibrium that is taxon specific (e.g., von Grafenstein et al., Reference von Grafenstein, Erlernkeuser and Trimborn1999; Decrouy, Reference Decrouy, Horne, Holmes, Rodriguez-Lazaro and Viehberg2012). For C. lacustris, the offset is well characterised as +1.2‰ ± 0.23 (von Grafenstein et al., Reference von Grafenstein, Erlernkeuser and Trimborn1999). For Ilyocypris, the other genus analysed here, there is no published offset. Moreover, we do not have sufficient numbers of paired analyses from Hoxne to be able to estimate the effect with confidence. However, values for paired analyses of Candona neglecta, which has a well-established vital offset, and I. bradyi in Belis and Aritztegui (Reference Belis and Ariztegui2004) have a mean difference of −1.78 ± 0.80 (n = 5), indicating that the vital offset for the latter is smaller than that for the former. We used a bootstrap technique (Efron and Tibsharini, Reference Efron and Tibsharini1993) to estimate the mean of the vital effect for Ilyocypris as +0.41‰ ± 0.31. The fact that the δ18O values for C. lacustris are more positive than those for Ilyocypris for the two levels at Hoxne for which we have values for both species is at least consistent with this result.
Table 9. Carbon and oxygen stable isotope results (‰) from Stratum C ostracod shells, Hoxne.a

The carbon isotope composition of lacustrine carbonates is strongly controlled by the δ13C of dissolved inorganic carbon (DIC): fractionation of carbonate isotopes between carbonate and DIC is minimally affected by temperature and, for ostracod shells, appears to be an equilibrium process (Keatings et al., Reference Keatings, Heaton and Holmes2002). The isotopic composition of DIC is, in turn, controlled by the δ13C of groundwater inputs, exchange with atmospheric CO2, and processes occurring in the lake, especially the preferential uptake of 12C during aquatic photosynthesis and the return of 13C-depleted organic matter from plants as a result of decay (Talbot, Reference Talbot1990).
The δ18O values for Ilyocypris fall within a fairly narrow range of −2.47‰ to −1.08‰, which equates to −2.88‰ to −1.49‰ at oxygen isotope equilibrium. Owing to taxonomic uncertainties and limited information on life cycles, it is difficult to be precise about the calcification season of the Ilyocypris specimens that were analysed. However, given that reconstructed temperatures were lower than present day temperatures during the deposition of Stratum C, especially during winter, it seems reasonable to assume that the ostracods were more likely to have reached adulthood during the warm season: this is supported for Ilyocypris sp. by limited evidence from Lake Geneva (Decrouy, Reference Decrouy2009). If this is correct, the calculated δ18O for Hoxne lake water during Stratum C times (based on reconstructed July temperature of 14°C) would have been −2.3‰, substantially higher than precipitation. Present-day precipitation of eastern England has a δ18O value of around −7‰ (Darling et al., Reference Darling, Bath and Talbot2003), and values during the colder climate of Stratum C times may have been even lower. Higher reconstructed δ18O values for the Hoxne Lake are best explained by evaporative enrichment. The limited number of δ18O values for C. lacustris provides partial support for these conclusions (Fig. 5, Table 9). Two of the levels yield similar results to those derived from analyses of Ilyocypris, assuming warm-season calcification, whereas for the remaining two, values are much more negative. However, even for the most negative values, reconstructed δ18O values for lake water (around −5.5‰) are higher than those for precipitation. Whether these values reflect variations in the lake environment during the formation of Stratum C remains unclear. Our assumption that temperatures remained constant during this interval is also questionable in view of the MOTR evidence for variation. However, substantially lower temperatures would be required for the ostracods to have calcified in water with a δ18O value close to the likely values of contemporary rainfall. We therefore conclude that Hoxne lake water was subject to evaporative enrichment throughout much or all of the time represented by Stratum C.
The carbon isotope values are more difficult to interpret, as they are controlled by a number of variables, as discussed earlier, and we have limited constraint on any one of these. Our conclusions must therefore remain speculative. Carbon supply to DIC in the Hoxne lake could have come from chalk, soil CO2 (ultimately derived from catchment vegetation), and aquatic vegetation, potentially modified by aquatic photosynthesis and equilibration with atmospheric CO2. Carbon in groundwater would have δ13C value in the range −16‰ to −10.5‰, assuming a 50:50 mixture of carbon from chalky till (δ13C for Upper Chalk Formation = +2‰ to +3‰: Jarvis et al., Reference Jarvis, Gale, Jenkyns and Pearce2006) and soil organic matter (δ13C = −24‰ to −24‰ in areas of C3 vegetation: Smith and Epstein, Reference Smith and Epstein1971; Deines, Reference Deines, Fritz and Fontes1980). Ostracod shells that calcified in equilibrium with this DIC would have δ13C values between −15‰ and −9.5‰. However, ostracods from Hoxne Stratum C have δ13C values between −3.1‰ and −5.7‰, indicating that other sources of carbon and/or processes must have affected DIC composition. The addition of 13C-depleted carbon to the DIC pool from respiration or decay of aquatic plants would decrease the δ13C of DIC. However, the uptake of 12C by aquatic plants for photosynthesis and equilibration with atmospheric CO2 both have the potential to increase δ13C values (e.g., Talbot, Reference Talbot1990). Without further evidence, it is difficult to be certain which was the dominant process, although partial equilibration with atmospheric CO2, which is a slow process, is consistent with a relatively long residence time for the lake, a condition that would favour evaporative enrichment of water isotopes, which is consistent with the oxygen isotope values and some of the faunal assemblage data.
MPC palaeotemperature reconstructions
The acronyms used in this section are explained in the “Methods,” but a summary look-up table is provided for convenience (Table 10). The MPC approach produced MORs for stratum C (July) and Strata D, E, and F (all January and July), while Stratum C (January) required acceptance of the only MGR result. Only a single proxy method (MOTR) was available for Stratum B, and no proxies usable for quantitative palaeotemperature reconstruction were available from Strata A and G. These results are presented here in chronological order from bottom to top and illustrated in Figure 4. For comparison, present-day mean monthly air temperature values for the vicinity of Hoxne are +3.0°C for January and +16.5°C for July.
Table 10. Look-up table of method acronyms used in the multiproxy consensus palaeotemperature reconstructions section of the text.
Stratum F
Overlap of the January BMCR and MOTR ranges gives an MOR of −8°C to −4°C. The BMCR and MOTR ranges together produced a July MOR of +16°C (i.e., a single value rather than a range), consistent with plant threshold values. July mean air temperature was thus very close to that of today, but January was at least 7.0°C colder, consistent with the suggestion (West, Reference West1956; Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008) that lake deposition began in the Anglian late-glacial period.
Stratum E
No BMCR could be obtained from Stratum E, but the CTF results for July fall within wider ranges indicated by the MOTR method, with an MOR of +18.0°C to +20.5°C (rounding up the upper and lower extremes to the nearest half degree). This is consistent with the evidence of three plant taxa that require a mean July temperature of +13.0°C or higher and of two beetle taxa (R. elongatus and X. rufovillosum) that indicate warmer than present-day climate, suggesting a full interglacial climate with summers between 1.5°C and 4.0°C warmer than today. Mean January air temperatures in Stratum E were similar to those of today or up to 3°C colder, as indicated by the 0°C to +3.0°C MOR of the MOTR results combined with two plant indicators.
Stratum D
The January MOR of the BMCR and plant thresholds is 0°C to +1°C, indicating winters that were 2°C to 3°C colder than those of today. The MOR result for July (+16.0°C to +19.0°C) is based on the BMCR method and plant indicator taxa, consistent with an interglacial climate, with summer temperatures not dissimilar to those of today, but possibly up to 2.5°C warmer.
Hiatus
A missing interval of unknown (but substantial) duration is represented by the erosion surface at the base of Stratum C, to the extent that pollen zones HoIIIb and HoIV are absent (Turner, Reference Turner1970; Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008).
Stratum C
The January reconstruction uses a combined BMCR overlapping the MOTR to produce an MOR of −7°C to −2°C, indicating winters between 5°C and 10°C colder than today. However, accommodating the two floral indicator taxa, which have minimum thresholds higher than the upper limit of the MOR by 0.5°C (H. helix) and 2.0°C (I. aquifolium), results in an MGR of −2°C to 0°C, indicating winters between 3°C and 5°C colder than today. This gap is accommodated within the variability apparent in the high-resolution record from Stratum C (discussed below). Whether the MOR or the MGR is accepted, the indications are clear that Stratum C summers and winters were definitely colder than during the interglacial period. The upper limit of the combined BMCR for July coincides with the lower limit of the MOTR, producing an MOR of precisely +14°C, indicating summers 2.5°C cooler than today. All three CTF ranges and the floral indicator taxa for Stratum C are consistent with this result.
In the other sedimentary units, it has necessarily been assumed that the sampled interval is representative of the unit as a whole, but in the case of Stratum C, high-resolution sampling has produced a detailed ostracod record that reveals variability within the unit. Reconstructed July palaeotemperature ranges from beetles and chironomids (neither of which were sampled at the same high resolution as the ostracods) show some overlap and some gaps when mapped onto the detailed MOTR results (Fig. 5, Table 11). The gaps may be the result of a lack of exact equivalence (both geographic within the Hoxne site and stratigraphic within Stratum C) in sampling for the different proxies, in view of the possibility of climate variation within this interval, as shown by the high-resolution MOTR results. We cannot rule out the possibility that at least some of the variation in the MOTR results may be due to “noise” attributable to low numbers of taxa recovered from some samples (the maximum number of species used for the MOTR analysis in a single sample was nine, but in a few cases only one or two could be used and inevitably produced very wide ranges), nor can we completely rule out contamination of the assemblages with reworked specimens. The BMCR results, although based on samples mostly of coarser stratigraphic resolution than the MOTR data and covering only parts of the succession, similarly show variation within Stratum C. However, the July results from the BMCR and CTF methods show a general correspondence with the minimum curve of the July MOTR results; similarly the January BMCR results mostly fall around the minima of the January MOTR results, although there are gaps in the lowest samples, the BMCR method indicating colder temperatures than the MOTR method. This suggests that the January and July MOTR minima may offer the best representation of the actual palaeotemperatures. The occurrences of the extinct ostracod I. quinculminata, which we consider to be possibly indicative of continental climate with warmer summers and colder winters than today, correspond to some extent with some of the warmer July intervals with cooler January temperatures.
Table 11. Stratum C individual proxy-based palaeotemperature reconstructions and multiproxy consensus (MOR in bold, MGR in italics) results.a

The maxima of the January BMCR results generally fall substantially below the threshold temperature indicated by pollen taxa H. helix and I. aquifolium, but in one case (a beetle sample equivalent to ostracod samples 343–345), the BMCR maximum matches the pollen taxa thresholds. Unfortunately, we have no way of determining precisely from West's (Reference West1956) pollen data the equivalence of his Stratum C samples with ours; his pollen diagrams for Stratum C (West, Reference West1956, figs. 22–24) show sporadic rather than continuous occurrences of the two pollen taxa. Molluscs and vertebrates recovered from Stratum C are generally consistent with the quantitative reconstructions; Dicrostonyx is indicative of tundra and mammoth steppe, while the temperature ranges of the suggested modern analog region for the fish assemblage has a July maximum that matches our July MOR, and although its January maximum is 6°C colder than the low end of our January MGR, this is easily accommodated within the variability shown by the high-resolution MOTR and BMCR results.
Stratum B
Only wide-ranging MOTR results are available for Stratum B (January −8°C to +13°C, July +10°C to +25°C), not inconsistent with the climatic amelioration suggested by mammalian and fish fauna (Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008), but also not ruling out the possibility of a significantly warmer or colder climate.
Stratum A
No palaeotemperature reconstructions are available for Stratum A, but its vertebrate assemblage suggests a transition from a warm to a cold climate (Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008).
DISCUSSION
Reworking in Stratum C assemblages
Discrepancies between proxy-based palaeotemperature reconstructions at some levels could be explained by reworking of certain “warm” taxa from exposed marginal deposits of the larger interglacial lake into the smaller Stratum C lake, an issue already discussed in the individual proxy results sections. It is possible that some Stratum C thermophilic chironomids were reworked from earlier warm period sediments; we consider this unlikely for well-represented temperate taxa that were not also found in Stratum E assemblages, but we cannot rule out the possibility that they were derived from material subsequently eroded and represented by the hiatus. The Stratum C beetle and pollen assemblages contain both warm and cold elements, suggestive of an admixture of material eroded from older interglacial deposits, but the beetle assemblages can be interpreted as in situ rather than contaminated with reworked material. Some degree of reworking of ostracods cannot be ruled out but is not required to explain the results, due to the 100% overlap of taxa used in the MOTR reconstructions. The vertebrate assemblage is consistent with a cold climate. While reworking of some vertebrate fossils from older deposits cannot be ruled out, taphonomic considerations suggest that dominant fish taxa represent an indigenous fauna, as does the presence of lemming, which may be regarded as indicating severe climatic conditions, at least during the deposition of the upper part of Stratum C. There are no obligate cold-climate molluscan taxa, but no component of the molluscan assemblage is incompatible with cold climate; the absence of some species in the underlying lacustrine part of the sequence identifies them as in situ elements of the Stratum C fauna. The gastropod B. tentaculata, usually found only in interglacial and interstadial deposits in Britain, has usually been considered a probable reworked element of Stratum C assemblages. However, its in situ presence is not necessarily ruled out by its modern distribution, as it has been recorded in Scandinavia at least as far north as the Gulf of Bothnia and Finland as well as northwestern Russia in Karelia and the area between St. Petersburg and Archangelsk (Alexandrowicz, Reference Alexandrowicz1999). Present-day mean January and July air temperatures in this region are between −13°C to −4°C and +14°C to +17°C, respectively, ranges that are not inconsistent with our MPC results for Stratum C.
Combined ostracod palaeoecology and oxygen stable isotope analyses confirm the Stratum C early cold phase and allow the tentative suggestion that warmer and/or more evaporatively enriched conditions prevailed at the beginning and end of the interval represented by these sediments; this is consistent with the evidence of the sample-by-sample BMCR results. Taking account of sedimentologic and stratigraphic evidence for reworking of older lake strata into Stratum C, West (Reference West1956, p. 334) considered that the pollen in Stratum C is “to a large extent derived from the older deposits, as must be many of the macrofossils.” If this is the case, we might be justified in ignoring our MGR for Stratum C January, which is necessitated by two pollen taxa (with assumed species-level identifications) and accepting the MOR result that shows winters 5°C to 10°C colder than those of today. However, as already discussed earlier for various proxies, invoking reworking is more a matter of interpretation than of hard evidence. West argued that the contemporary flora of Stratum C were characteristic of an arctic or subarctic climate, with Salix herbacea, Salix myrsinites, S. polaris, and B. nana growing around the lake. Betula nana indicates mean July air temperature of +7°C or higher. According to West's view, we should probably consider the persistent presence (but low abundance) of T. latifolia in Stratum C to be the result of its reworking from older lake sediments exposed around the margins during an interval of lower lake level; certainly, it is relatively abundant in parts of Stratum D. However, if considered as an in situ component of the lake flora during Stratum C times, T. latifolia indicates mean July air temperatures of at least +13°C, consistent with the July MOR of +14°C and with the indications of the small vertebrate assemblage (warmest month between +7°C and +14°C).
Comparisons with other sites and records
Ashton et al. (Reference Ashton, Lewis, Parfitt, Penkman and Coope2008) suggested that the Hoxne climate intervals could be correlated with those evident in the SPECtral MApping Project (SPECMAP) stack (e.g., Tzedakis et al., Reference Tzedakis, Andrie, de Beaulieu, Birks, Crowhurst, Follieri and Hooghiemstra2001): the Hoxnian interglacial with MIS 11c, the “Arctic Bed”/Stratum C stadial with MIS 11b, and the Stratum B interstadial with MIS 11a. They noted, however, that other correlations might be possible, for example, with isotopic events seen in more detailed deep-ocean records. Multiproxy evidence from deep Atlantic Ocean sediment cores has shown that the inception of the MIS 10 glacial period was complex, with a series of cold/warm climatic oscillations following the MIS 11 interglacial period (Desprat et al., Reference Desprat, Sánchez Goñi, Turon, McManus, Loutre, Duprat, Malaize, Peyron and Peypouquet2005; Martrat et al., Reference Martrat, Grimalt, Shackleton, de Abreu, Hutterli and Stocker2007; Voelker et al., Reference Voelker, Rodrigues, Billups, Oppo, McManus, Stein, Hefter and Grimalt2010; Rodrigues et al., Reference Rodrigues, Voelker, Grimalt, Abrantes and Naughton2011; Candy et al., Reference Candy, Schreve, Sherriff and Tye2014). Off the Iberian Peninsula, these are evident in the terrestrial pollen record and pollen-derived air temperature reconstructions (Desprat et al., Reference Desprat, Sánchez Goñi, Turon, McManus, Loutre, Duprat, Malaize, Peyron and Peypouquet2005), alkenone-derived sea surface temperatures (Rodrigues et al., Reference Rodrigues, Voelker, Grimalt, Abrantes and Naughton2011), and planktonic foraminiferal δ18O and lithic abundance interpreted primarily as ice-rafted debris (Voelker et al., Reference Voelker, Rodrigues, Billups, Oppo, McManus, Stein, Hefter and Grimalt2010). Similar climatic oscillations are evident in the Lake Baikal biogenic silica record (Prokopenko et al., Reference Prokopenko, Bezrukova, Khursevich, Solotchina, Kuzmin and Tarasov2010).
There is disagreement in the literature as to how the marine isotope substages and isotopic events should be correlated (Candy et al., Reference Candy, Schreve, Sherriff and Tye2014). Some authors, with reference to SPECMAP, have considered the possibility that the MIS 11b stadial and MIS 11a interstadial equate to events 11.22 and 11.1, respectively (e.g., Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008), while others have matched MIS 11b with 11.24 and then place 11.23–11.1 in MIS 11a (e.g., Voelker et al., Reference Voelker, Rodrigues, Billups, Oppo, McManus, Stein, Hefter and Grimalt2010). Desprat et al. (Reference Desprat, Sánchez Goñi, Turon, McManus, Loutre, Duprat, Malaize, Peyron and Peypouquet2005) identified four post-interglacial cold intervals within MIS 11: the first (oldest) they equated with isotopic event 11.24, the second forms a short cold episode within the 11.23 interstadial, while the third was assigned to 11.22. Subsequently Voelker et al. (Reference Voelker, Rodrigues, Billups, Oppo, McManus, Stein, Hefter and Grimalt2010, fig. 6) identified the same four cold intervals as stadials (numbered I–IV in decreasing order of age) separated by three interstadials; the first (oldest) stadial was placed in MIS 11b, which they equated with isotopic event 11.24, separating the 11.23 interstadial from the MIS 11c interglacial period, while the remaining three stadials were assigned to MIS 11a. Thus, MIS 11b has been centred around 375 ka BP (e.g., SPECMAP stack: Imbrie et al., Reference Imbrie, Hays, Martinson, McIntyre, Mix, Morley, Pisias, Prell, Shackleton, Berger, Imbrie, Hays, Kukla and Saltzman1984) or around 390 ka BP (e.g., Voelker et al., Reference Voelker, Rodrigues, Billups, Oppo, McManus, Stein, Hefter and Grimalt2010).
Figure 8 shows a possible correlation of our Hoxne palaeotemperature reconstructions with examples of the abovementioned palaeoclimate records (locations shown in Fig. 9). We cannot tell how long an interval is represented by the hiatus between Stratum D and Stratum C at Hoxne. The erosion of strata deposited between the lowest (remaining) part of Stratum D and the beginning of Stratum C might have removed a record of climate oscillations that is now represented only by material that has been reworked, mixed, and incorporated into Stratum C or removed entirely from the basin; this could have included not only the late part of the interglacial but also post-interglacial cold and/or warm intervals. Nevertheless, although the possibility of contamination of Stratum C assemblages with reworked material cannot be entirely ruled out, the simplest solution is to correlate Stratum C/Stratum B with the first post-interglacial cold/warm oscillation of Voelker et al. (Reference Voelker, Rodrigues, Billups, Oppo, McManus, Stein, Hefter and Grimalt2010), 11.24/11.23. Human occupation at Hoxne therefore likely occurred during the 11.23 interstadial, represented by Stratum B. It is unfortunate that our reconstructed palaeotemperature ranges for this interval are wide and inconclusive, allowing for both summers and winters to have been colder or warmer than those of the present day; perhaps the evidence of a boreal forest suggests a greater likelihood that temperatures were nearer the cooler ends of the reconstructed ranges, although this may be difficult to reconcile with the mammal and fish evidence of climatic amelioration (Ashton et al., Reference Ashton, Lewis, Parfitt, Penkman and Coope2008). Another possibility is that Stratum B represents a warm 11.23 interval punctuated by a short cold interval, as is evident in the pollen record off the Iberian Peninsula (Desprat et al., Reference Desprat, Sánchez Goñi, Turon, McManus, Loutre, Duprat, Malaize, Peyron and Peypouquet2005) and the Lake Baikal biogenic silica record (Prokopenko et al., Reference Prokopenko, Bezrukova, Khursevich, Solotchina, Kuzmin and Tarasov2010). The lesson of Stratum C, discussed earlier, may be pertinent: the integration of proxy evidence into a single virtual sample representative of the whole unit, blurring any record of such a short climate oscillation, could explain the wide MOTR results and other conflicting evidence. Similarly, the lack of high-resolution continuous sampling through Stratum E at Hoxne precludes any attempt to recognise an abrupt cold-climate episode within MIS 11c, identified at Marks Tey in Essex and ODP 980 in the North Atlantic by Candy et al. (Reference Candy, Tye, Coxon, Hardiman, Matthews and Palmer2021), who correlated the two occurrences by means of a single tephra layer. On the other hand, dynamic fluvial depositional environments are generally far less capable of preserving high-resolution archives of environmental change than are lacustrine depositional settings, so even high-resolution sampling of Stratum B might be unlikely to yield any record of such short-term changes.

Figure 8. Comparison of records of MIS 11 climatic variability. From left to right: (A) Structure and subdivision of MIS 11 in the LR04 benthonic δ18O stack (Lisiecki and Raymo, Reference Lisiecki and Raymo2005) and the planktonic δ18O Low Latitude Stack (Bassinot et al., Reference Bassinot, Labeyrie, Vincent, Quidelleur, Shackleton and Lancelot1994); (B) Lake Baikal BDP-96 composite record of biogenic silica, a proxy for lacustrine productivity (Prokopenko et al., Reference Prokopenko, Bezrukova, Khursevich, Solotchina, Kuzmin and Tarasov2010); (C) pollen record of temperate and humid trees from North Atlantic oceanic core MD01-2447 off northwestern Iberia (Desprat et al., Reference Desprat, Sánchez Goñi, Turon, McManus, Loutre, Duprat, Malaize, Peyron and Peypouquet2005); (D) alkenone-derived sea-surface temperature from North Atlantic oceanic core MD03-2699 off western Iberia (Rodrigues et al., Reference Rodrigues, Voelker, Grimalt, Abrantes and Naughton2011) and pollen-derived mean air temperature of coldest (MTCO) and warmest (MTWA) months from North Atlantic oceanic core MD01-2447 off northwestern Iberia (Desprat et al., Reference Desprat, Sánchez Goñi, Turon, McManus, Loutre, Duprat, Malaize, Peyron and Peypouquet2005); (E) Hoxne pollen zones, strata, and reconstructed January and July air temperature ranges (this study). Site locations are shown in Fig. 9.

Figure 9. Map showing locations of sites in Fig. 8.
Two other British terrestrial MIS 11 sites may record post-interglacial cold/warm oscillations that could be correlatives of the Stratum C/Stratum B interval at Hoxne. One is Quinton in the West Midlands, where a cold interlude was recognised in BMCR results, although the pollen record at the site shows no equivalent response (Coope and Kenward, Reference Coope and Kenward2007). The other is West Stow in Suffolk, where the lower part of the sequence represents the early part of the interglacial period, while the upper part (Bed 7), separated from the lower interval by a hiatus, contains cold-climate indicators (including ostracods, molluscs, and mammals) as well as thermophilous mammals and fish (Preece et al., Reference Preece, Parfitt, Bridgland, Lewis, Rowe and Atkinson2007), the mixture being suggestive of a similar situation to that discussed earlier for Stratum B at Hoxne. The MOTR results for Bed 7 at West Stow (January −7°C to −1°C, July +15°C to +21°C) suggest a more continental climate, with winters at least 4°C colder than today, and are quite similar to those for Stratum C at Hoxne (Benardout, Reference Benardout2015). It is possible that the cold intervals at the different sites each correlate with a different isotopic episode in the deep-ocean record, but it is worth considering that, given their occurrence within a relatively small geographic area, it is not unreasonable to expect that all three experienced the same climatic variations capable of influencing their sedimentary and fossil records in similar ways. The similarities among these three British records thus hint at equivalence and a common cause reflecting a regional climatic oscillation worthy of further investigation at other British MIS 11 sites, for example, Marks Tey in Essex, where there is a substantial lacustrine succession overlying the Hoxnian interglacial deposits (Horne et al., Reference Horne, Bal, Benardout, Huckstepp, Lewis, March, Bridgland, Allen and White2014; Tye et al., Reference Tye, Sherriff, Candy, Coxon, Palmer, McClymont and Schreve2016).
CONCLUSIONS
The multiproxy comparisons of BMCR, MOTR, and CTF results, combined with threshold temperatures for indicator taxa, produce consistent and well-constrained palaeotemperature ranges with few disagreements.
The results confirm and constrain two post-Anglian temperate phases at Hoxne (the first represented by Strata F, E, and D, being the Hoxnian interglacial period; the second represented by Strata B and A) separated by a cold episode (Stratum C), tentatively correlated with MIS 11 substages. Multiproxy consensus palaeotemperature reconstructions (°C), for January and July, respectively, were as follows: Stratum F, −8°C to −4°C and +16°C; Stratum E, 0°C to +3.0°C and +18.0°C to +20.5°C; Stratum D, 0°C to +1°C and +16.0°C to +19.0°C; Stratum C, −2°C to 0°C and +14°C. For Stratum B, a single-proxy reconstruction of January −8°C to +13°C and July +10°C to +25°C was obtained; no reconstruction was possible for Stratum A.
Discrepancies between proxy methods may be explicable in terms of (1) local reworking of older (interglacial Strata E/D) deposits into the younger (Stratum C) lake, (2) lack of precisely equivalent samples for different proxies, and (3) differences in sampling resolution between the proxies.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/qua.2022.20
Acknowledgments
The data, interpretation, and discussion presented herein began as a contribution to the Ancient Human Occupation of Britain project (funded by the Leverhulme Trust) and is completed as a contribution to the Pathways to Ancient Britain project (funded by the Calleva Foundation). The Hoxne Dating Project (HDP) was funded by AHOB, and SAP is grateful to Glenys Salter and Robert Symmons for their expert processing of the finds from the HDP excavations. SJB is grateful to Pat Haynes for picking and slide mounting the chironomid samples. We thank the two reviewers, Jim Rose and Ian Boomer, as well as associate editor Vera Aldeias, for their constructive suggestions, which helped us to improve the article. We have no competing interests to declare.
Author contributions
DJH: coordination, editing, preparation of figures, MOTR reconstructions, pollen and plant macrofossil palaeoclimate indications. NA: site context and stratigraphy. GB: ostracod analyses and interpretation, MOTR reconstructions. SJB: chironomid analyses and interpretation, CTF reconstructions. GRC: beetle analyses and interpretation, MCR reconstructions. JAH: isotope analyses and interpretation. SGL: site context and stratigraphy. SAP: site context and stratigraphy, vertebrate analyses and interpretation. TSW: mollusc analyses and interpretation. NJW: beetle interpretation, MCR reconstructions. JEW: ostracod analyses and interpretation, MOTR reconstructions. All authors contributed to the multiproxy consensus reconstructions as well as discussion of reworking and comparisons with other sites and records.

























