



Introduction
A helminthic infectious disease, schistosomiasis affects 240 million people worldwide and has spread to 74 nations and regions. It is most common in tropical and sub-tropical areas (WHO 2022). After the eggs mechanically occlude the microvasculature, early pathological alterations appear, leading to acute vasculitis with endothelial injury and necrosis (Van de Vijver et al. Reference Van de Vijver KK, Jacobs, Kuypers, Hokke, Deelder and Van Marck2006). Following the enrollment of immunological and inflammatory cells at infection areas because of the lodged ova’s sustained antigenic stimulation, some infected people develop chronic fibrosis and granulomas (Chuah et al. Reference Chuah, Jones, Burke, McManus and Gobert2014). For liver fibrosis in schistosomiasis, activation of hepatic stellate cells is essential (Zhang et al. Reference Zhang, Fang and Liu2019). Interleukin-1β (IL-1β), tumor necrosis factor (TNF), and transforming growth factor (TGF) are a few examples of inflammatory cytokines or chemokines that Kupffer cells (KCs) can swiftly release in response to hepatic damage (Wree et al. Reference Wree, McGeough, Peña, Schlattjan, Li, Inzaugarat, Messer, Canbay, Hoffman and Feldstein2014). These elements may promote hepatic stellate cell (HSC) activation and proliferation, which may lead to the production of extracellular matrix (ECM), and so cause liver fibrosis (Wree et al. Reference Wree, McGeough, Inzaugarat, Eguchi, Schuster, Johnson, Peña, Geisler, Papouchado, Hoffman and Feldstein2018).
Nucleotide-binding oligomerization domain-like (NOD-like) receptors are intracellular sensors of pathogen-associated molecular patterns that are linked to cell stress and damage-associated chemical patterns that enter the cell by phagocytosis or holes. They are important players in the control of the innate immune response (Mahla et al. Reference Mahla, Reddy, Prasad and Kumar2013). Toll-like receptors and NOD-like receptors can work together to regulate inflammatory and apoptotic responses (Franchi et al. Reference Franchi, Warner, Viani and Nuñez2009). Complex protein NOD-like receptor protein 3 (NLRP3) is primarily produced in macrophages and is a part of the inflammasome that recognizes by-products of injured cells (Lu and Wu Reference Lu and Wu2015). Caspase-1 activation can encourage the generation of IL-1β and IL-18 and protect against certain pathogen damage (Cao et al. Reference Cao, Wang, Long and He2019). In situations including Angiotensin II, steatohepatitis induced by alcohol, and drug-induced liver damage, there is growing evidence connecting the inflammation caused by the NLRP3 inflammasome leading to tissue damage and liver fibrosis (Nagy Reference Nagy2015; Ning et al. Reference Ning, Luo, Wang, Li, Pan, Yang, Ling, Huang, Ma, Jin, Wang and Li2017). According to earlier research, the NLRP3 inflammasome regulates macrophage and HSC activity, which is essential for the advancement of fibrosis (Inzaugarat et al. Reference Inzaugarat, Johnson, Holtmann, McGeough, Trautwein, Papouchado, Schwabe, Hoffman, Wree and Feldstein2019; Wree et al. Reference Wree, McGeough, Inzaugarat, Eguchi, Schuster, Johnson, Peña, Geisler, Papouchado, Hoffman and Feldstein2018; Zhang et al. Reference Zhang, Fang and Liu2019). New therapeutic drugs for treating hepatic fibrosis offer a promising target in the inhibition of the NLRP3 inflammasome.
Autophagy is a cellular mechanism that has been conserved throughout evolution and is crucial for the survival and upkeep of cells by destroying organelles, proteins, and macromolecules (Ding et al. Reference Ding, Fu, Wang, Liu, Wang and Wu2021). Pro-IL-1β, mitochondria, and the inflammasome components are all degraded by autophagosomes, which reduces IL-1β release (Levine and Deretic Reference Levine and Deretic2007). In response to metabolic or extrinsic stress, NLRP3 inflammasome activation was enhanced in macrophages with impaired autophagy (Lee et al. Reference Lee, Kim, Quan, Lee, Kim, Kim, Bae, Hur and Lee2016).
There are seven sirtuins (SIRT1 to 7) in mammalian cells. Sirtuins are a class of nicotinamide adenine dinucleotide+ (NAD+) dependent deacetylases. Sirtuin 3 (SIRT3) is largely located in mitochondria as the major regulator of mitochondrial protein acetylation (Xu et al. Reference Xu, Hertzel, Steen and Bernlohr2016). SIRT3 is a key regulator of mitochondrial functions that control energy production, metabolism, and the body’s reaction to oxidative stress. It is highly expressed in tissues with a high rate of metabolic activity, including the liver (Hirschey et al. Reference Hirschey, Shimazu, Goetzman, Jing, Schwer, Lombard, Grueter, Harris, Biddinger, Ilkayeva, Stevens, Li, Saha, Ruderman, Bain, Newgard, Farese, Alt, Kahn and and Verdin2010). Additionally, SIRT3-induced autophagy suppressed NLRP3 inflammasome activation in macrophages (Liu et al. Reference Liu, Huang, Wei, Gao, Huang, Sun, Zhu and Shen2018).
Although chemotherapy successfully eradicates adult worms and inhibits the accumulation of schistosome eggs, less effective medications are focused on treating hepatic fibrosis that has already developed, particularly in the chronic and severe phases of the illness. In recent years, significant efforts have been made to find medications that target hepatic fibrosis in schistosomiasis (Niu et al. Reference Niu, Hu, Hong, Li and Shen2022).
As an anti-rheumatic medication, auranofin has been used in therapeutic settings. It stops osteoclast differentiation by inhibiting the NLRP3 inflammasome. (Kuntz et al. Reference Kuntz, Davioud-Charvet, Sayed, Califf, Dessolin, Arnér and Williams2007). Additionally, the mechanism by which auranofin inhibits schistosomiasis has been thoroughly studied (Angelucci et al. Reference Angelucci, Sayed, Williams, Boumis, Brunori, Dimastrogiovanni, Miele, Pauly and Bellelli2009). It was found that auranofin reduced worm burden in S. mansoni-infected mice likely through inhibition of Thioredoxin glutathione reductase (TGR) (Kuntz et al. Reference Kuntz, Davioud-Charvet, Sayed, Califf, Dessolin, Arnér and Williams2007), and it has in vitro worm-killing action (Kim et al. Reference Kim, Kim, Kim, Kim, Lee and Kang2019). Two experimental liver fibrosis models were used to evaluate the possibility of using auranofin clinically to treat liver fibrosis (Kim et al. Reference Kim, Choi, Kim, Kim, Jun, Yoon, Kim, Hwang, Kim and Lim2021); however, the liver fibrosis caused by S. mansoni differs from other models of liver fibrosis in that the host is impacted by the stimulation of the eggs and the growth of the worm. Therefore, we hypothesized that auranofin’s anti-inflammatory effects, activating macrophage autophagy and preventing inflammasome activation, will alleviate S. mansoni egg-induced liver granuloma and later development of hepatic fibrosis.
Material and methods
Animals
Eighty male BALB/c laboratory-bred mice were purchased from Theodor Bilharz Research Institute (TBRI), Imbaba, Giza, Egypt. They were eight weeks old, weighed 18–20 grams, and were parasite-free. After receiving the approval of the National Liver Institute’s Ethical Committee (IRB00383/2022), they were maintained at 27±2°C. They were kept to a standard diet and water ad libitum. The cages were cleaned twice a week to ensure good sanitary conditions. This was done in compliance with the ARRIVE guidelines (Percie du Sert et al. Reference Percie du Sert, Hurst, Ahluwalia, Alam, Avey, Baker, Browne, Clark, Cuthill, Dirnagl, Emerson, Garner, Holgate, Howells, Karp, Lazic, Lidster, MacCallum, Macleod, Pearl, Petersen, Rawle, Reynolds, Rooney, Sena, Silberberg, Steckler and Würbel2020).
Cercariae
Cercariae of a virulent strain of S. mansoni were isolated from infected Biomphelaria alexandrina snails raised and maintained at TBRI. The S. mansoni cercariae were isolated by the procedure described in Liang et al. (Reference Liang, Bruce and Boyd1987), and the removed cercariae were counted by a stereomicroscope (Mohamed et al. Reference Mohamed, El-Din, Mohamed and Habib2011). Mice were infected subcutaneously in a dose of 60±10 cercariae/mouse using the technique in Holanda et al. (Reference Holanda, Pellegrino and Gazzinelli1974).
Drug and dose
Auranofin® was provided by Tocris Bioscience (a Bio-Techne Brand, Minneapolis, Minnesota, USA) and was dissolved in 100% ethanol. A dose of auranofin that is safe for healthy mice was given orally at a dose of 1 mg/kg twice daily for nine days (Andrade et al. Reference Andrade, Chaparro, Capparelli and Reed2014; Debnath et al. Reference Debnath, Ndao and Reed2013).
Experimental design
The mice were divided into four groups each consisting of 20 mice. (1) Negative control group (Group I/GI); non-infected non-treated mice which were subdivided equally into two subgroups (GIa and GIb) each including 10 mice. (2) Positive control group (Group II/GII); infected non-treated mice which were subdivided equally into two subgroups (GIIa and GIIb) each including 10 mice. (3) Early treated group (Group III/GIII) infected mice were treated orally by gastric gavage with auranofin (1 mg/kg twice daily for 9 days), starting on the 4th week post-infection (p.i.). (4) Late-treated group (Group IV/GIV) infected mice were treated orally by gastric gavage with auranofin (1 mg/kg twice daily for 9 days) beginning the 6th week p.i. (This helps in analyzing auranofin’s impact on S. mansoni egg-induced liver fibrosis) (Table 1).
Table 1. Experimental groups

The mice were divided into four groups, each consisting of 20 mice.
(1) Negative control group (Group I/GI): non-infected, non-treated mice which were subdivided equally into two subgroups (GIa and GIb), each including 10 mice.
(2) Positive control group (Group II/GII): infected, non-treated mice which were subdivided equally into two subgroups (GIIa and GIIb), each including 10 mice.
(3) Early-treated group (Group III/GIII): infected mice were treated orally by gastric gavage with auranofin (1 mg/kg twice daily for 9 days), starting on the 4th week post-infection (p.i.).
(4) Late-treated group (Group IV/GIV): infected mice were treated orally by gastric gavage with auranofin (1 mg/kg twice daily for 9 days), beginning the 6th week p.i. (This helps in analyzing auranofin’s impact on S. mansoni egg-induced liver fibrosis).
N: number per group
Sacrifice and perfusion
Mice from groups GIa, GIIa, and GIII were sacrificed on the 6th week p.i. (anesthetized with ether and euthanized by decapitation) to assess the effect of auranofin on S. mansoni egg-induced liver granuloma. From each mouse, portions of the liver were taken after the worm count to determine the egg burden and oogram pattern. For histopathological studies, liver samples were collected. Sera were isolated from blood samples and used for biochemical investigations. To determine how auranofin affects S. mansoni-induced liver fibrosis; mice of the GIb, GIIb, and GIV groups were sacrificed on the 10th week p.i., and liver samples were collected for histopathological and immunohistochemical studies. As with the early-treated group, blood samples were also collected, and sera were divided for biochemical investigations.
Parasitological studies
Porto-mesenteric worm burden and reduction %
S. mansoni adult worms (males, females, and couples) that emerged from the portomesenteric system following perfusion were counted. The percentage reduction of adult worms after treatment was calculated according to Tendler et al. (Reference Tendler, Pinto, Lima, Gebara and Katz1986) using the formula R = C−V/C × 100, where R = reduction %, C = mean number of adult worms from infected non-treated mice, and V = mean number of parasites from treated mice.
Tissue egg load/g intestine and liver
The livers of sacrificed infected mice were weighed and then digested in 5% KOH solution. The number of eggs per gram of liver was calculated under a light microscope at 40× magnification (Cheever Reference Cheever1969).
Oogram pattern
Three fragments from the ileum of each mouse (1 cm each) were cut longitudinally, rinsed with saline, and dried gently using Whatman filter paper. They were examined microscopically, and the percentage of different egg developmental stages (oogram pattern) was studied using the technique of Pellegrino et al. (Reference Pellegrino, Oliveira, Faria and Cunha1962), which involved identifying and counting eggs at various stages of maturity.
Histopathological studies
Specimens from the liver tissue of mice infected with S. mansoni and treated with auranofin were obtained. The specimens were preserved in 10% neutral buffered formalin (pH 7.2), embedded in paraffin blocks, and sliced at a thickness of 4–5 μm. The tissue sections from all groups were stained according to Bancroft and Gamble (Reference Bancroft and Gamble2008). The histopathological characteristics were evaluated using standard H&E to determine the number and measure the diameter of Schistosoma granulomas using twenty non-overlapping high-power fields (400X). Eosinophils per granuloma region were also counted. Moreover, types of granulomas were assessed and divided into four major evolutionary stages: (1) pre-granulomatous exudative, characterized by an infiltrate of inflammatory cells in the process of organization around the parasite egg; (2) necrotic-exudative, distinguished by a central halo of necrosis and numerous inflammatory cells dispersed irregularly on subsequent layers; (3) exudative-productive, distinguished by a rich structure of collagen fibers and inflammatory cells concentrated in the periphery and exhibiting a more organized and circumferential aspect; and (4) productive, with a characteristic thick band of collagen fibers separating the egg from a small number of inflammatory cells. The total area connected to the granulomatous reaction was used to calculate the size of granulomas using semi-automated methods (Amaral et al. Reference Amaral, Silva, Dias, Malta, Rosa, Costa-Neto, Gentile and Melo2017). In addition, Masson trichome (MT) stain was done to assess collagen deposition in the liver tissue of groups GIIb and GIV. Briefly, slides were stained for 10 minutes with Weigert’s iron hematoxylin working solution, washed, stained for 15 minutes with Biebrich scarlet-acid fuchsin solution, washed, immersed for 15 minutes in phosphomolybdic-phosphotungstic acid solution, transferred directly to aniline blue solution for 10 minutes, quickly washed in distilled water, and then transferred to 1% acetic acid solution for 5 minutes. Collagen strands were stained blue, while hepatocytes were stained red. Microscopy and photo taking were carried out using an Olympus CX41 microscope and a DP26 camera in Shinjuku, Tokyo, Japan.
Immunohistochemical (IHC) studies
The streptavidin-biotin system was used for the immunostaining procedure. Interleukin-1β (IL-1β) diluted to 1:50 (Cat. #12242), Sirtuin 3 (SIRT3) diluted to 1:200 (Cat. #2627), and smooth muscle actin (SMA) diluted to 1:400 (Cat. #19245) were obtained from Cell Signaling Technology, Massachusetts, USA. Before antigen retrieval, the tissue was deparaffinized and rehydrated in a high pH ethylenediaminetetraacetic acid (EDTA) solution for 20 minutes. After that, it was cooled at room temperature for an additional 20 minutes. The primary antibodies were applied to each plate and left overnight at 4°C. Horseradish peroxidase/3,3’-diaminobenzidine (HRP/DAB) secondary antibody, ready for applying Mayer’s hematoxylin as a counterstain and DAB chromogen substrate to visualize the staining, SIRT3, IL-1β, and SMA markers were employed. In every test, both positive and negative controls were used. SMA and IL-1β antibodies exhibited cytoplasmic localization, whereas SIRT3 antibody showed nuclear and cytoplasmic localization. Histoscore (H score) was used to evaluate expression. The intensity and percentage of cells were multiplied as follows: (strong intensity (3) x percentage) + (moderate intensity (2) x percentage) + (mild intensity (1) x percentage) + (negative staining (0) x percentage). A final score between 0 and 300 was established (Pu et al. Reference Pu, Storr, Zhang, Rakha, Green, Ellis and Martin2017).
Assessment of liver function
Serum samples were separated by centrifugation at 3000 rpm for ten minutes. Liver function tests for alanine transaminase (ALT) and aspartate transaminase (AST) were performed using an Integra 400 autoanalyzer (Roche, Germany). Quantitative determination of AST and ALT was carried out by the Warburg method with pyridoxal-5-phosphate at a wavelength of 340 nm according to the International Federation of Clinical Chemistry (IFCC) protocol (Bergmeyer et al. Reference Bergmeyer, Horder and Rej1986).
Statistical analysis
Data were collected, tabulated, and analyzed by Statistical Package of Social Science (SPSS) version 22 (Armonk, NY: IBM Corp, 2013) and GraphPad Prism version 8.0.0 for Windows (GraphPad Software, San Diego, California, USA). The mean (X) and standard deviation (SD) were used to present quantitative data, and the T test was used to compare data between two groups. Numbers and percentages were used to show qualitative data. To compare qualitative variables, the Monto-Carlo test was used. Statistics were considered significant at P values ≤ 0.05.
Results
The overall S. mansoni worm burden and ova gram/liver tissue were significantly decreased (63.07% and 78.26%, respectively, P < 0.001) in GIII when compared to the infected control group (GIIb), whereas in GIV, the decrease was 56.08% and 69.67%, respectively (Table 2). Regarding the schistosomal oogram pattern, there was a statistically significant difference between the means of immature eggs and mature ova (26.80 ± 1.25 and 20.40 ± 0.872, respectively, P < 0.001) in the early-treated group (GIII) and the control infected group (GIIa), which were (55.20 ± 2.20 and 43.40 ± 1.50, respectively, P < 0.001). In addition, there was a significant difference between the two groups regarding the mean number of dead eggs, which was 38.70 ± 1.44 in the GIII group against 4.20 ± 0.86 in the control-infected group (P < 0.001). Meanwhile, in GIV, the means of immature eggs and mature ova were 29.70 ± 1.44 and 24.24 ± 0.65, respectively, and the mean number of dead eggs was 40.50 ± 1.69 with a statistically significant difference with GIIb (P < 0.001) (Table 3).
Table 2. Comparison of mean S. mansoni worm burdens and egg load studied groups

T test was used. Values are expressed as mean ± standard deviation. GIIa: early positive control group (n=10); GIII: early-treated group infected mice with auranofin (n=20), ♂: male, ♀: female; GIIb: late positive control group (n=10); GIV: late-treated group infected mice with auranofin (n=20), ♂: male, ♀: female
Table 3. Comparison of oogram patterns in studied groups

T test was used. Values are expressed as mean ± standard deviation. GIIa: early positive control group (n=10); GIII: early-treated group infected mice with auranofin (n=20); GIIb: late positive control group (n=10); GIV: late-treated group infected mice with auranofin (n=20)
Histopathological results
There was a high density of hepatic granuloma and a significantly larger area of eosinophil-rich granuloma in the infected control subgroups (GIIa and GIIb) (Figure 1a and d and Table 4). While in the early-treated GIII, the reduction percent in granuloma number and size was 40.63% (P < 0.001) and 48.66% (P = 0.108), respectively, with a predominant exudative/necrotic granuloma type and reduced eosinophilic count (Figure 1b and e and Table 4). Meanwhile, the best reduction in granuloma number and size was 76.63% (P < 0.001) and 67.08% (P = 0.008), respectively, in GIV (the late-treated group) (Table 4), and the form of granuloma was productive, with a low eosinophil count (Figure 1c and f). The infected and treated groups both had Schistosoma granulomas at various phases of development.
Table 4. The comparison between different groups regarding the pathological data

A T test was used to compare mean ± SD (standard deviation); the Monto-Carlo test was used to compare qualitative data. GIIa and GIIb: early and late positive control group (n=10); GIII: early-treated group infected mice with auranofin (n=20); GIV: late-treated group with praziquantel and auranofin (n=20)
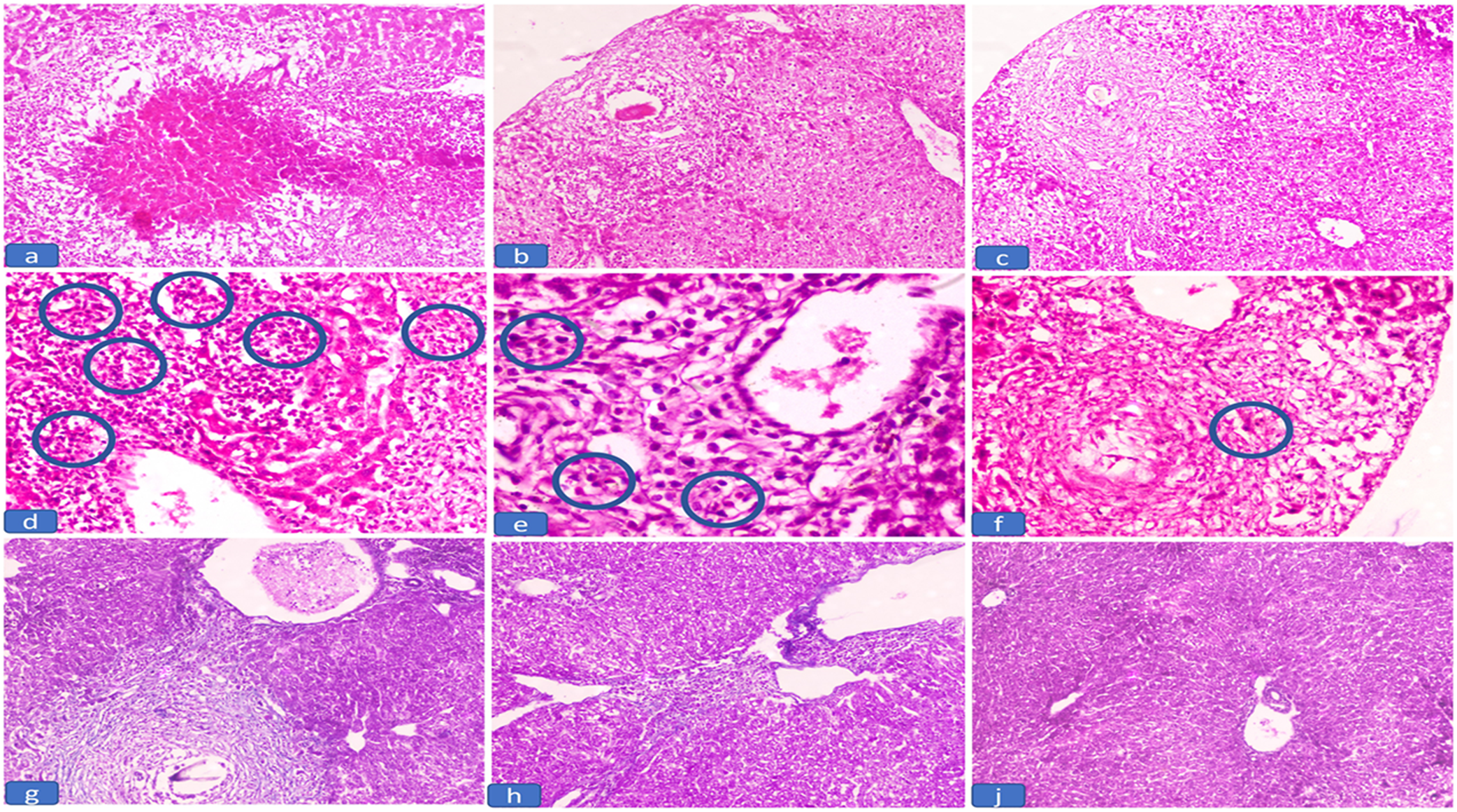
Figure 1. The histopathological findings of liver tissue in the studied groups. a) Exudative necrotic granuloma in infected untreated groups (GIIa and GIIb) (H&E, 100x), b) Exudative granuloma in early-treated group (GIII) (H&E, 100x), c) Productive granuloma in the late-treated group (GIV) (H&E, 100x), d) Dense eosinophilic count in the inflammatory infiltrate in the infected untreated groups (GIIa and GIIb) (circles) (H&E, 400x), e) Decrease eosinophilic count in the inflammatory in the early-treated groups (GIII) (circles) (H&E, 200x), f) Few eosinophilic count in the inflammatory in the late-treated groups (GIV) (circles) (H&E, 200x), g) Marked periportal fibrosis in infected untreated group (GIIb) (MT, 100x), h) Mild periportal fibrosis in the late-treated group (GIV) (MT, 100x), j) No periportal fibrosis in the late-treated group (GIV) (MT, 100x).
Collagen deposition in the liver tissue assessed by MT stain was significantly decreased in the GIV (late-treated group) as compared to the GIIb (infected control) that showed marked periportal fibrosis (Figure 1g, h, and j and Table 4).
When compared to the infected groups, the density of hepatic granulomas and the eosinophilic-rich infiltrate were significantly lower in both treated groups (P < 0.001) (Table 4). In GIV, the granulomas were significantly smaller than those in the late-infected group (P = 0.008). In addition, the degree of fibrosis was significantly decreased (P = 0.025) (Table 4).
Immunohistochemical results
Interleukin-1β expression was significantly reduced in the GIII and GIV, with GIV significantly exhibiting the lowest expression (P < 0.001), whereas the infected control groups (GIIa and GIIb) had significantly higher levels of IL-1β (H score was 82.00 ± 4.89 and 94.00 ± 2.44, respectively) (Table 5 and Figure 2a, b, and c). However, SIRT3 expression was low in GIIa and GIIb (H score was 14.00 ± 2.91 and 13.00 ± 4.35, respectively). SIRT3 expression was significantly higher in both treated groups compared to the infected, with the highest expression observed in GIV (late-treated) (H score was 224.00 ± 5.61) (P < 0.001) (Table 5 and Figure 2d, e, and f). The liver tissue of GIIa and GIIb revealed significant SMA expression in both portal tracts and sinusoidal HSC (H score was 69.00 ± 2.00 and 83 ± 3.00, respectively). Smooth muscle actin expression was highly significantly reduced in GIV (H score was 10.50 ± 1.38) (P < 0.001) (Table 5 and Figure 2g, h, and j).
Table 5. Comparative expression of IL-1β, SIRT3, and SMA during the early and late phases of infected and treated groups

IL-1β: interleukin 1β, SIRT3: Sirtuin 3, SMA: Smooth muscle actin, HSC: Hepatic stellate cells. T test was used, mean ± SD (standard deviation). GIIa and GIIb: early and late positive control group (n=10); GIII: early-treated group infected mice with auranofin (n=20); GIV: late-treated group with auranofin (n=20)

Figure 2. The immunohistochemical expression of IL-1β, SIRT3, and SMA in the studied groups. a) Mice infected untreated group (GIIa and GIIb) showed high IL-1β H score expression in the portal tracts (IHC, 100x), b) Mice early-treated group (GIII) showed moderate IL-1β H score expression in the portal tracts (IHC, 100x), c) Mice of late-treated group (GIV) showed low IL-1β H score expression in the portal tracts (IHC, 100x), d) Mice of infected untreated group (GIIa and GIIb) showed low SIRT3 H score expression in the portal tracts (IHC, 100x), e) Mice of early-treated group (GIII) showed moderate SIRT3 H score expression in the portal tracts (IHC, 100x), f) Mice of late-treated group (GIV) showed high SIRT3 H score expression in the portal tracts (IHC, 100x), g) Mice of infected untreated late group (GIIb) showed high SMA H score expression in the portal tracts (IHC, 100x), h) Mice of late-treated group (GIV) showed mild SMA H score expression in the portal tracts (IHC, 100x), j) Mice of late-treated group (GIV) showed mild SMA H score expression in the portal tracts (IHC, 100x).
Results of ALT and AST serum levels
The serum levels of ALT and AST were improved in both treated groups, GIII (31.76 ± 0.30 and 45.73 ± 0.76) and GIV (28.25 ± 0.36 and 39.22 ± 0.31), respectively (P < 0.05), when compared to the infected control group, with a significant difference, GIIa (65.88 ± 0.32 and 76.88 ± 0.68) and GIIb (79.02 ± 0.52 and 89.28 ± 0.41), respectively (P < 0.05). Additionally, there was a notable difference between the two treated groups, GIII and GIV (Figure 3).

Figure 3. Alanine aminotransferase serum and aspartate aminotransferase serum levels IU/L (mean ± SD) in the different groups studied. GIa and GIb: early and late negative control group each (n=10); GIIa and GIIb: early and late positive control group each (n=10); GIII: early-treated group infected mice with auranofin (n=20); GIV: late-treated group auranofin (n=20).
ALT: alanine aminotransferase, AST: aspartate aminotransferase
*Mean values were significantly different from the GIa group: P < 0.05
**Mean values were significantly different from the GIb group: P < 0.05
#Mean values were significantly different from the GIIa group: P < 0.05
##Mean values were significantly different from the GIIb group: P < 0.05
Discussion
The most severe pathological manifestations of schistosomiasis are hepatic granulomas and fibrosis, which are caused by egg deposition in the liver (Zhang et al. Reference Zhang, Fang and Liu2019). The development of hepatic fibrosis is heavily influenced by chronic inflammation and the activation of inflammasomes (Alegre et al. Reference Alegre, Pelegrin and Feldstein2017). Auranofin is clinically approved for the treatment of rheumatoid arthritis, and it has lately been researched for potential therapeutic uses in a variety of illnesses (Chen et al. Reference Chen, Chao, Huang and Fang2022; Steers et al. Reference Steers, Chen, O’Leary, Du, Van Beek and Cullen2022). In this work, we examined the impact of auranofin on liver granuloma and fibrosis caused by S. mansoni eggs.
The administration of auranofin to infected mice resulted in a significant decrease in the total worm burden and tissue egg load with an increase in the degenerating ova, contrary to the infected control group. Auranofin decreased worm burden in mice with S. mansoni infection, probably via inhibiting TGR (Kuntz et al. Reference Kuntz, Davioud-Charvet, Sayed, Califf, Dessolin, Arnér and Williams2007).
Regarding the histopathological results, when compared to the control groups, auranofin considerably reduced the mean hepatic granuloma diameter and number in both early and late treatment groups (P < 0.001), with a predominance of degraded ova. Masson’s trichrome staining showed that auranofin reduced fibrosis and significantly improved gross hepatic morphology with a lower percentage of collagen in the late-treated group. Since there are no first-line treatments for liver fibrosis, a study’s findings demonstrate auranofin therapeutic efficacy in liver fibrosis (Kim et al. Reference Kim, Choi, Kim, Kim, Jun, Yoon, Kim, Hwang, Kim and Lim2021).
In this study, the S. mansoni-infected group had significantly higher serum levels of ALT and AST, which indicated liver damage, as previously reported by El-Refai et al. (Reference El-Refai, Atia and Mahmoud2019). Administration of auranofin significantly improved their levels in both treated groups. Similar results were reported by Kim et al. (2023) who stated that mice treated with auracyanide had normal serum levels of AST and ALT, indicating that auracyanide, an active metabolite of auranofin, may prevent liver fibrosis.
NLRP3 inflammasome has been the focus of some of the most recent research to date regarding its effects on numerous human diseases, antiviral responses, and antifungal responses (He et al. Reference He, Chen, Zhu, An, Dong, Yu, Zhang, Wu, Li and Zhang2018; Lee et al. Reference Lee, Kim, Kim, Shong, Ku and Jo2013; Mao et al. Reference Mao, Zhang, Li, Chen, Wang, Wu, Guo, Lu, Yang and An2014; Vakrakou et al. Reference Vakrakou, Boiu, Ziakas, Xingi, Boleti and Manoussakis2018). Additionally, it can be found in the cytoplasm of HSCs and can be triggered by several different pathogens, including S. mansoni, E. coli, and S. japonicum (Chen et al. Reference Chen, Cheng, Chang, Cao, Feng, Chen, Lam and Peng2019; Meng et al. Reference Meng, Xia, Lu, Wang, Boini, Li and Tang2016). The activation of caspase-1 and the subsequent release of IL-1β are made possible by the NLRP3 inflammasome, which is crucial in the development of hepatic fibrosis and inflammation (Kaufmann et al. Reference Kaufmann, Kui, Reca, Leszczynska, Kim, Booshehri, Wree, Friess, Hartmann and Broderick2022). In addition, collagen deposition and NLRP3 activation in primary human hepatocytes results in pyroptotic cell death (Gaul et al. Reference Gaul, Leszczynska, Alegre, Kaufmann, Johnson, Adams, Wree, Damm, Seehofer and Calvente2021). The adaptive innate immune responses to the development of fibrosis are controlled by the NLRP3 inflammasome. In liver fibrosis, IL-1β and TNF-α promote interactions between immune cells and HSCs (Seki and Schwabe Reference Seki and Schwabe2015).
We examined auranofin’s impact on the release of IL-1β to clarify how it affects the NLRP3 inflammasome’s activation. This study demonstrated that auranofin therapy decreased IL-1β expression in the GIII (early) and GIV (late) treated groups, with GIV significantly exhibiting the lowest expression. It demonstrates that auranofin limits the synthesis of inflammatory cytokines and inhibits innate immune signaling by acting on numerous domains. Similarly, Zhang et al. (Reference Zhang, Chen, Gao, Hou, Gu, Gui, Huang, Liu, Ren and Wang2012) discovered that hepatocyte necrosis and granulomatous inflammation are both down-regulated by reduced expression of IL-6 and IL-1β (pro-inflammatory cytokines) (Zhang et al. Reference Zhang, Chen, Gao, Hou, Gu, Gui, Huang, Liu, Ren and Wang2012). Blocking the NLRP3 inflammasome was the underlying mechanism and thus reduced the release of IL-1β and decreased the HSCs’ capacity to migrate. Auranofin has an anti-inflammatory impact, and we hypothesized that inhibiting inflammasome activation would contribute to this effect and markedly decrease the region of granuloma and fibrosis in the liver in the current investigation.
Auranofin did not affect the NLRP3 inflammasome’s priming signals. It only prevented caspase-1 from cleaving and IL-1β from maturing. Due to thioredoxin reductase modulation, previous studies have found increased levels of intracellular reactive oxygen species and decreased release of IL-1β by auranofin (Isakov et al. Reference Isakov, Weisman-Shomer and Benhar2014; Lee et al. Reference Lee, Xu, Chiu, Leibold, Tse, Bao, Yuen, Chan, Lai and Chin2019). Similarly, Hwangbo et al. (Reference Hwangbo, Kim, Ji, Kim, Lee, Kim, Park, Keum, Hong and Cheong2020) revealed that auranofin may have the potential to be a candidate for reducing the symptoms of non-alcoholic fatty liver disease by inhibiting the NLRP3 inflammasome (Hwangbo et al. Reference Hwangbo, Kim, Ji, Kim, Lee, Kim, Park, Keum, Hong and Cheong2020). Auranofin was also shown by Kim et al. (Reference Kim, Choi, Kim, Kim, Jun, Yoon, Kim, Hwang, Kim and Lim2021) to considerably slow the progression of fibrosis in a thioacetamide- or carbon tetrachloride-induced fibrotic liver (Kim et al. Reference Kim, Choi, Kim, Kim, Jun, Yoon, Kim, Hwang, Kim and Lim2021). Additionally, Kaufmann et al. (Reference Kaufmann, Kui, Reca, Leszczynska, Kim, Booshehri, Wree, Friess, Hartmann and Broderick2022) found that the production of pro-inflammatory and pro-fibrotic cytokines as well as the onset of liver inflammation and fibrosis depends on the activation of the NLRP3 inflammasome complex in myeloid cells (Kaufmann et al. Reference Kaufmann, Kui, Reca, Leszczynska, Kim, Booshehri, Wree, Friess, Hartmann and Broderick2022). Production of cytokines, such as IL-1β, may occur after the activation of the NLRP3 inflammasome in KCs. In mice infected with S. japonicum, this activates HSCs and is accompanied by NLRP3 inflammasome activation, which is linked to the development of hepatic fibrosis (Zhang et al. Reference Zhang, Fang and Liu2019).
The present study showed that α-SMA expression in both portal tracts and sinusoidal HSC was significantly reduced in the auranofin late-treated group (GIV). Auranofin inhibits the migration of HSCs, which are the main suppliers of collagen and are essential for hepatic fibrogenesis. Quiescent HSCs are activated and transdifferentiated into myofibroblast-like cells under pathological circumstances; these cells express the myofibroblast marker and α-SMA and have increased collagen production (Xu et al. Reference Xu, Hertzel, Steen and Bernlohr2016). Consequently, one of the key features of HSC activation has been the development of α-SMA, which has grown in significance as a measure of hepatic fibrosis (Tsuchida and Friedman Reference Tsuchida and Friedman2017).
Sirtuin 3 has beneficial effects on cellular responses and is sensitive to metabolic state (Liu et al. Reference Liu, Huang, Wei, Gao, Huang, Sun, Zhu and Shen2018). Here, we investigated how SIRT3-mediated autophagy affects NLRP3 inflammasome activity. The present study showed auranofin treatment causes a significantly higher expression of SIRT3. Similarly, Mukherjee et al. (Reference Mukherjee, Mo, Paolella, Perry, Toth, Hugo, Chu, Tong, Chellappa and Baur2021) established that the mitochondrial sirtuin, SIRT3, is crucial for liver regeneration (Mukherjee et al. Reference Mukherjee, Mo, Paolella, Perry, Toth, Hugo, Chu, Tong, Chellappa and Baur2021). SIRT3 may help prevent liver illnesses such as non-alcoholic fatty liver, liver damage, and fibrosis, according to mounting evidence (Xu et al. Reference Xu, Zhu, Bai, Xia, Li, Lu, Li and Gao2019). Similar results were obtained by Wang et al. (Reference Wang, Li, Gu, Chen, Duanmu, Miao, Yao, Tao, Tu, Xiong, Zhao and Liu2020), who discovered that SIRT3 activation reduced liver fibrosis.
Loss of SIRT3 causes animals to be more sensitive to ischemia and other injuries (Lombard et al. Reference Lombard, Alt, Cheng, Bunkenborg, Streeper, Mostoslavsky, Kim, Yancopoulos, Valenzuela, Murphy, Yang, Chen, Hirschey, Bronson, Haigis, Guarente, Farese, Weissman, Verdin and and Schwer2007), and Sirtuin 3 has emerged as the main isoform responsible for reducing mitochondrial proteins (Zhang et al. Reference Zhang, Xiang, Liu, Chen, He and Liu2020a). Furthermore, Zhang et al. (Reference Zhang, Fang, Linghu, Liu, Gan and Lin2020b) reported that AEDC (12βacetoxy-16,23-epoxy-24,25-dihydroxy3β-9,19-cyclolanost-22(23)-ene) inhibited the NLRP3 inflammasome through SIRT3-autophagy, which may be used to treat metabolic disorders linked to adipose tissue inflammation (Zhang et al. Reference Zhang, Fang, Linghu, Liu, Gan and Lin2020b).
Autophagy-mediated NLRP3 inflammasome inhibition was concomitant with increases in SIRT3 levels (Liu et al. 2017), indicating that it might act as a mediator for auranofin’s liver regeneration effects (Liu et al. Reference Liu, Huang, Wei, Gao, Huang, Sun, Zhu and Shen2018). Furthermore, Liu et al. (2017) demonstrated that SIRT3-deficient macrophages showed increased NLRP3 inflammasome activation, endothelial dysfunction, and defective autophagy (Liu et al. Reference Liu, Huang, Wei, Gao, Huang, Sun, Zhu and Shen2018). For the majority of liver cell types, autophagy is a cytoprotective and anti-fibrotic process, and it is essential for maintaining the metabolic homeostasis of hepatocytes (Khambu et al. Reference Khambu, Yan, Huda, Liu and Yin2018). Numerous disorders have been linked to the interaction between autophagy and the NLRP3 inflammasome, and the activation of this enzyme that causes pyroptosis has been linked to inflammation, fibrosis, and cell death in the liver (Al Mamun et al. Reference Al Mamun, Akter, Hossain, Sarker, Safa, Mustafa, Muhammad and Munir2020). Similarly, Wang et al. (Reference Wang, Ren, Yuan, Ma, Shi and Ding2018) and Hao et al. (Reference Hao, Cui, Gao, Wang, Liu, Li, Hou and Jia2022) demonstrated that lowering the levels of NLRP3, IL-1, IL-18, and caspase-1 improved hepatic dysfunction (Hao et al. Reference Hao, Cui, Gao, Wang, Liu, Li, Hou and Jia2022; Wang et al. Reference Wang, Ren, Yuan, Ma, Shi and Ding2018). The production of inflammasome-dependent IL-1β is inhibited by macrophage autophagy, which reduces acute toxin-induced liver injury and death in a mouse model (Ilyas et al. Reference Ilyas, Zhao, Liu, Lin, Tesfa, Tanaka and Czaja2016). Alternatively, evidence indicates that autophagy plays a role in several anti-fibrotic processes that promote cell death and cell cycle arrest in HSCs or stop the production of pro-fibrotic mediators, such as excessive collagen accumulation (Lucantoni et al. Reference Lucantoni, Martínez-Cerezuela, Gruevska, Moragrega, Víctor, Esplugues, Blas-García and Apostolova2021).
Conclusion
Auranofin prevents hepatic granulomas and fibrosis brought on by S. mansoni eggs. The NLRP3 inflammasome was blocked as a component of the underlying mechanism and thus decreased the release of IL-1β. Moreover, autophagy-mediated NLRP3 inflammasome inhibition was concomitant with an increase in SIRT3 levels that mediate the effect of auranofin on liver regeneration. Together with the restoration of serum ALT and AST towards normal levels, these findings reveal how auranofin suppresses the development of liver fibrosis. Therefore, it is important to reconsider auranofin as a potential therapy for hepatic granuloma and subsequent fibrosis induced by S. mansoni eggs.
Data availability statement
The data is available upon reasonable request to the corresponding author.
Author contribution
Amany Fawzy Atia, Noha Mohamed Abou-Hussein, Dina Mohamed Sweed, Eman Sweed, and Noha Ahmed Abo-khalil made major contributions to the work presented, whether in the conception, study design, execution, data gathering, analysis, or interpretation. Furthermore, Amany Fawzy Atia, Noha Mohamed Abou-Hussein, Dina Mohamed Sweed, Eman Sweed, and Noha Ahmed Abo-khalil contributed to the article’s development and revision, agreed on the journal to which it will be submitted, and approved the final manuscript for publication. Finally, Amany Fawzy Atia, Noha Mohamed Abou-Hussein, Dina Mohamed Sweed, Eman Sweed, and Noha Ahmed Abo-khalil agreed to accept responsibility for all parts of the work, including ensuring that any doubts about the accuracy or integrity of any portion of the work are thoroughly explored.
Competing interest
No conflicts of interest were declared.
Ethics approval
The manuscript is exempt from the approval of the ethical committee.








