



Introduction
Biodiversity is critical to maintain functional ecosystems and healthy environments (Moranta et al. Reference Moranta, Torres, Murray, Hidalgo, Hinz and Gouraguine2021). In spite of this, the current large-scale loss of species and habitats is at an unprecedented rate and primarily driven by human activities. To halt negative trends of biodiversity loss, a wide array of conservation programs has been carried out worldwide. In this sense, long-term and well-planned conservation programmes represent the key to help halting the current extinction crisis (Ewen et al. Reference Ewen, Armstrong, Parker and Seddon2012). Among conservation efforts, conservation translocations (e.g. reintroduction and restocking; IUCN/SSC 2013) represent efficient tools to favour the recolonisation of part of the ancient range of a species and to boost the demographic growth of threatened populations, as well as their long-term conservation (Armstrong and Seddon Reference Armstrong and Seddon2008, Ewen et al. Reference Ewen, Armstrong, Parker and Seddon2012). Released individuals may come from captive-breeding programmes, be raised from eggs taken from clutches removed from wild populations (Colomer et al. Reference Colomer, Oliva-Vidal, Jiménez, Martínez and Margalida2020), or be translocated from natural donor source populations (Ewen et al. Reference Ewen, Armstrong, Parker and Seddon2012). For avian species, such interventions typically involve the release of individuals in areas where they were nearly extirpated or were supposed to live in the past (Carter et al. Reference Carter, Newbery, Grice and Hughes2008). Good results were achieved especially for projects targeting raptors, preventing population extinction, or favouring population increase. One of the most successful wildlife comeback stories is the reintroduction of the Bearded Vulture Gypaetus barbatus in the Alps (Schaub et al. Reference Schaub, Zink, Beissmann, Sarrazin and Arlettaz2009). Similar long-term conservation projects allowed the successful return of the osprey Pandion haliaetus (e.g. Dennis and Dixon Reference Dennis and Dixon2001, Monti et al. Reference Monti, Dominici, Choquet, Duriez, Sammuri and Sforzi2014) and of iconic scavengers such as the Eurasian Griffon Vulture Gyps fulvus and the Black Vulture Aegypius monachus (Slotta-Bachmayr et al. Reference Slotta-Bachmayr, Boegel and Camiña2004, Terrasse et al. Reference Terrasse, Sarrazin, Choisy, Clémente, Henriquet, Lécuyer, Pinna, Tessier, Chancellor and Meyburg2004) across Europe.
However, conservation translocations are a high-cost endeavour with a long history of failures and problems (Robert et al. Reference Robert, Colas, Guigon, Kerbiriou, Mihoub, Saint-Jalme and Sarrazin2015, Taylor et al. Reference Taylor, Canessa, Clarke, Ingwersen, Armstrong, Seddon and Ewen2017, Berger-Tal et al. Reference Berger-Tal, Blumstein and Swaisgood2020). Most common issues faced in such projects were: a) difficulties in removing local threats (Fasce and Fasce Reference Fasce and Fasce2009, Simón et al. Reference Simón, Couto, Lucio Carrasco, García–Baquero, Godino, Gutiérrez, Hernández, Jiménez, Liñán, López, Padial, Salamanca, Bautista, del Barco, Cabrera and and Macías2007); b) unsuitable regional or local conditions in contrast with movement strategies of the target species, such as the presence of ecological barriers affecting the survival of released birds (Phipps et al. Reference Phipps, López-López, Buechley, Oppel, Álvarez, Arkumarev, Bekmansurov, Berger-Tal, Bermejo, Bounas, Alanís, de la Puente, Dobrev, Duriez, Efrat, Fréchet, García, Galán, García-Ripollés, Gil, Iglesias-Lebrija, Jambas, Karyakin, Kobierzycki, Kret, Loercher, Monteiro, Morant Etxebarria, Nikolov, Pereira, Peške, Ponchon, Realinho, Saravia, Şekercioğlu, Skartsi, Tavares, Teodósio, Urios and Vallverdú2019, Buechley et al. Reference Buechley, Oppel, Efrat, Phipps, Carbonell Alanís, Álvarez, Andreotti, Arkumarev, Berger-Tal, Bermejo Bermejo, Bounas, Ceccolini, Cenerini, Dobrev, Duriez, García, García-Ripollés, Galán, Gil, Giraud, Hatzofe, Iglesias-Lebrija, Karyakin, Kobierzycki, Kret, Loercher, López-López, Miller, Mueller, Nikolov, de la Puente, Sapir, Saravia, Sekercioglu, Sillett, Tavares, Urios and Marra2021).
In this context, it has been claimed that the movement ecology of animals and of birds in particular, is a topic of paramount importance to consider when planning adequate conservation measures in space and time (Allen and Singh Reference Allen and Singh2016, Efrat et al. Reference Efrat, Hatzofe, Miller, Mueller, Sapir and Berger-Tal2022). This is especially true in reintroduction and restocking programs, where decisions on how, when and where releasing individuals into the wild can influence birds’ survival, and in turn, the success of the project itself (Schaub et al. Reference Schaub, Zink, Beissmann, Sarrazin and Arlettaz2009, Margalida et al. Reference Margalida, Colomer, Oro, Arlettaz and Donázar2015, Colomer et al. Reference Colomer, Oliva-Vidal, Jiménez, Martínez and Margalida2020). Mortality rate is widely assumed higher during migration than at other times of the year (Newton Reference Newton2007, Klaassen et al. Reference Klaassen, Hake, Strandberg, Koks, Trierweiler, Exo, Bairlein and Alerstam2014) and is often associated with the crossing of ecological barriers, which is energetically demanding and requires good body condition as well as advanced flight skills and experience (Sergio et al. Reference Sergio, Tanferna, De Stephanis, Jiménez, Blas, Tavecchia, Preatoni and Hiraldo2014). Indeed, mortality rate is known to be particularly high for juveniles during their first year of life because of their inexperience and reduced capacity to cope with hazards or unsuitable weather conditions encountered en route (Grande et al. Reference Grande, Serrano, Tavecchia, Carrete, Ceballos, Díaz-Delgado, Tella and Donázar2009, Sergio et al. 2011, De Pascalis et al. Reference De Pascalis, Panuccio, Bacaro and Monti2020).
To our knowledge, there are no specific studies dealing with the associated risks of migration posed by unsuitable weather conditions aimed at implementing the conservation translocation procedures within population restoration programmes. Thus, this research presents a novel and previously ignored important factor affecting the success of conservation translocations programmes targeting migratory bird species, especially large soaring raptors. Wildlife management and conservation may benefit by strengthening the link with movement ecology as knowledge on species movements can be used to improve the effectiveness of management actions and develop better conservation strategies (Allen and Singh Reference Allen and Singh2016, Margalida et al. Reference Margalida, Pérez-García, Afonso and Moreno-Opo2016).
In this paper, we discuss this topic by reviewing the available information related to the Egyptian Vulture Neophron percnopterus, a long-distance migratory raptor and one of the most endangered vulture species of Europe (Santangeli et al. Reference Santangeli, Girardello, Buechley, Botha, Di Minin and Moilanen2019, Keller et al. Reference Keller, Herrando, Voříšek, Franch, Kipson, Milanesi, Martí, Anton, Klvaňová, Kalyakin, Bauer and Foppen2020, Oppel et al. Reference Oppel, Arkumarev, Bakari, Dobrev, Saravia-Mullin, Solomon Adefolu, Aktay Sözüer, Tersoo Apeverga, Arslan, Barshep, Bino, Bounas, Çetin, Dayyoub, Dobrev, Duro, El-Moghrabi, El-Safoury, Endris, Ghazal Asswad, Hanson Harry, Ivande, Jbour, Kapsalis, Kret, Mahamued, Manu, Mengistu, Razack Moussa Zabeirou, Inuwa Muhammad, Nakev, Ngari, Onoja, Osta, Özuslu, Petrovski, Popgeorgiev, Pourchier, Ruffo, Shobrak, Sidiropoulos, Skartsi, Sözüer, Stara, Tesfaye, Topi, Vavylis, Velevski, Vorpsi, Wondafrash, Xeka, Yeniyurt, Yordanov and Nikolov2021a). In Europe the species has a patchy distribution, with Spain being one of the main strongholds hosting 32–47% of the estimated 3,195–4,832 European breeding pairs (Del Moral and Molina Reference Del Moral and Molina2018, Keller et al. Reference Keller, Herrando, Voříšek, Franch, Kipson, Milanesi, Martí, Anton, Klvaňová, Kalyakin, Bauer and Foppen2020). In Europe, it suffered a strong decline (>50%) over the last few decades (BirdLife International 2017) and has already disappeared from several countries, while many populations are continuing to decline (e.g. Sanz-Aguilar et al. Reference Sanz-Aguilar, Sanchez-Zapata, Carrete, Benitez, Avila, Arenas and Donázar2015). For these reasons, it has been considered as ‘Endangered’ by IUCN and it has been included in the Annex I of the "Birds Directive" of the European Union 70/409/EU. Egyptian Vultures are soaring migrants that rely primarily on thermal and orographic updrafts to gain altitude and glide to cover large distances. By migrating that way, this species needs to travel over land and tends to concentrate at geographical straits to avoid long flapping flights over the open sea, which represents an ecological barrier (Phipps et al. Reference Phipps, López-López, Buechley, Oppel, Álvarez, Arkumarev, Bekmansurov, Berger-Tal, Bermejo, Bounas, Alanís, de la Puente, Dobrev, Duriez, Efrat, Fréchet, García, Galán, García-Ripollés, Gil, Iglesias-Lebrija, Jambas, Karyakin, Kobierzycki, Kret, Loercher, Monteiro, Morant Etxebarria, Nikolov, Pereira, Peške, Ponchon, Realinho, Saravia, Şekercioğlu, Skartsi, Tavares, Teodósio, Urios and Vallverdú2019, Oppel et al. Reference Oppel, Buechley, López-López, Phipps, Arkumarev, Bounas, Williams, Dobrev, Dobrev, Stoychev, Kret, Cenerini, Ceccolini, Saravia, Nikolov, Panuccio, Mellone and Agostini2021b). At open sea, thermal currents supporting the soaring flight are known to be weaker than over land, especially in temperate zones (Bildstein Reference Bildstein2006, Duriez et al. Reference Duriez, Peron, Gremillet, Sforzi and Monti2018, but see Nourani et al. Reference Nourani, Bohrer, Becciu, Bierregaard, Duriez, Figuerola, Gangoso, Giokas, Higuchi, Kassara, Kulikova, Lecomte, Monti, Pokrowsky, Sforzi, Therrien, Tsiopelas, Vanstelaant, Viana, Yamaguchi, Wikelski and Kamran2021). Furthermore, unfavourable weather conditions are highly known to affect flight performance (Thorup et al. Reference Thorup, Alerstam, Hake and Kjellén2003, Oppel et al. Reference Oppel, Dobrev, Arkumarev, Saravia, Bounas, Kret, Velevski, Stoychev and Nikolov2015), potentially leading to mortality risk especially when crossing ecological barriers (Klaassen et al. Reference Klaassen, Hake, Strandberg, Koks, Trierweiler, Exo, Bairlein and Alerstam2014, Strandberg et al. Reference Strandberg, Klaassen, Hake and Alerstam2010).
In the Balkans, it has been speculated that causes of migration-related mortality in juvenile Egyptian Vultures may be an indirect consequence of low population numbers and decreasing trends (Oppel et al. Reference Oppel, Dobrev, Arkumarev, Saravia, Bounas, Kret, Velevski, Stoychev and Nikolov2015). Based on known data, the authors proposed that juveniles could potentially benefit from following experienced adult/immature birds guiding them along the route, despite there is no evidence that social facilitation is indispensable for successful migration (Oppel et al. Reference Oppel, Dobrev, Arkumarev, Saravia, Bounas, Kret, Velevski, Stoychev and Nikolov2015). Inexperienced juveniles raised in low density areas could accidentally choose to follow sub-optimal migratory routes, augmenting the risk of venturing into the open sea, with a possible consequent high risk of mortality.
In this regard, we hypothesize that the peculiarities in the movement capacity of the species and the opportunities for meeting migrating conspecifics, together with the regional or local wind regimes, could influence the choice of the migratory route to follow, with possible repercussions on the survival of inexperienced juvenile Egyptian Vultures. This could be remarkable in geographical areas facing wide water bodies, where birds have to cope with long sea-crossings during migration, such as the case of Southern Italy, where a relict Egyptian Vulture population exists and has been the subject of a captive-breeding restocking programme since 2003 (Ceccolini and Cenerini Reference Ceccolini and Cenerini2016, Oppel et al. Reference Oppel, Buechley, López-López, Phipps, Arkumarev, Bounas, Williams, Dobrev, Dobrev, Stoychev, Kret, Cenerini, Ceccolini, Saravia, Nikolov, Panuccio, Mellone and Agostini2021b).
To verify this hypothesis, we analyse and report on the prevailing wind patterns along the main migratory route used by Italian Egyptian Vultures to cross the Central Mediterranean, during both spring and autumn, and coupled this information with count data of migrating Egyptian Vultures, as per field observations carried out over Marettimo Island (Egadi Archipelago). We also compared and commented on this information with published data from other Egyptian Vulture populations. Finally, we discuss 1) how these concomitant factors could further influence the long-term survival of the Italian Egyptian Vulture population, already critically affected by anthropogenic factors, and 2) how taking into account both environmental and geographic characteristics could ultimately help to take more effective management decisions and planning conservation strategies.
Movement ecology and mortality across flyways
The Egyptian Vulture is a highly philopatric long-lived raptor species, with sex-biased dispersal and delayed maturity (Grande et al. Reference Grande, Serrano, Tavecchia, Carrete, Ceballos, Díaz-Delgado, Tella and Donázar2009, Serrano et al. Reference Serrano, Cortés-Avizanda, Zuberogoitia, Blanco, Benítez, Ponchon, Grande, Ceballos, Morant, Arrondo, Zabla, Montelío, Ávila, González, Arroyo, Frías, Kobierzycki, Arenas, Tella and Donázar2021). It is a partial migratory species with both migratory and resident populations across its broad distribution range encompassing three continents (Ferguson-Lee and Christie Reference Ferguson-Lees and Christie2001, Keller et al. Reference Keller, Herrando, Voříšek, Franch, Kipson, Milanesi, Martí, Anton, Klvaňová, Kalyakin, Bauer and Foppen2020, Oppel et al. Reference Oppel, Buechley, López-López, Phipps, Arkumarev, Bounas, Williams, Dobrev, Dobrev, Stoychev, Kret, Cenerini, Ceccolini, Saravia, Nikolov, Panuccio, Mellone and Agostini2021b). Migratory populations in the Western Palearctic follow three major flyways (Fig. 1a): i) a western flyway, followed by individuals breeding in Southern France and the Iberian Peninsula across the Strait of Gibraltar and towards wintering areas in West Africa. This journey usually takes 2–3 weeks and covers c.3,500 km (Oppel et al. Reference Oppel, Buechley, López-López, Phipps, Arkumarev, Bounas, Williams, Dobrev, Dobrev, Stoychev, Kret, Cenerini, Ceccolini, Saravia, Nikolov, Panuccio, Mellone and Agostini2021b); ii) an eastern flyway, followed by individuals breeding in the Balkans and Turkey, which migrate around the eastern coast of the Mediterranean towards the Red Sea and winter in eastern sub-Saharan Africa. This journey usually takes 3–4 weeks and covers a distance of more than 5.000 km (Oppel et al. Reference Oppel, Arkumarev, Bakari, Dobrev, Saravia-Mullin, Solomon Adefolu, Aktay Sözüer, Tersoo Apeverga, Arslan, Barshep, Bino, Bounas, Çetin, Dayyoub, Dobrev, Duro, El-Moghrabi, El-Safoury, Endris, Ghazal Asswad, Hanson Harry, Ivande, Jbour, Kapsalis, Kret, Mahamued, Manu, Mengistu, Razack Moussa Zabeirou, Inuwa Muhammad, Nakev, Ngari, Onoja, Osta, Özuslu, Petrovski, Popgeorgiev, Pourchier, Ruffo, Shobrak, Sidiropoulos, Skartsi, Sözüer, Stara, Tesfaye, Topi, Vavylis, Velevski, Vorpsi, Wondafrash, Xeka, Yeniyurt, Yordanov and Nikolov2021a; Fig. 1a); and iii) a central flyway, followed by individuals of the relict Italian population (Ceccolini and Cenerini Reference Ceccolini and Cenerini2016; Fig. 1b). This journey can take 3–4 weeks and covers a distance of about 4,000 km (Ceccolini et al. Reference Ceccolini, Cenerini and Aebischer2009, www.lifegyptianvulture.it).
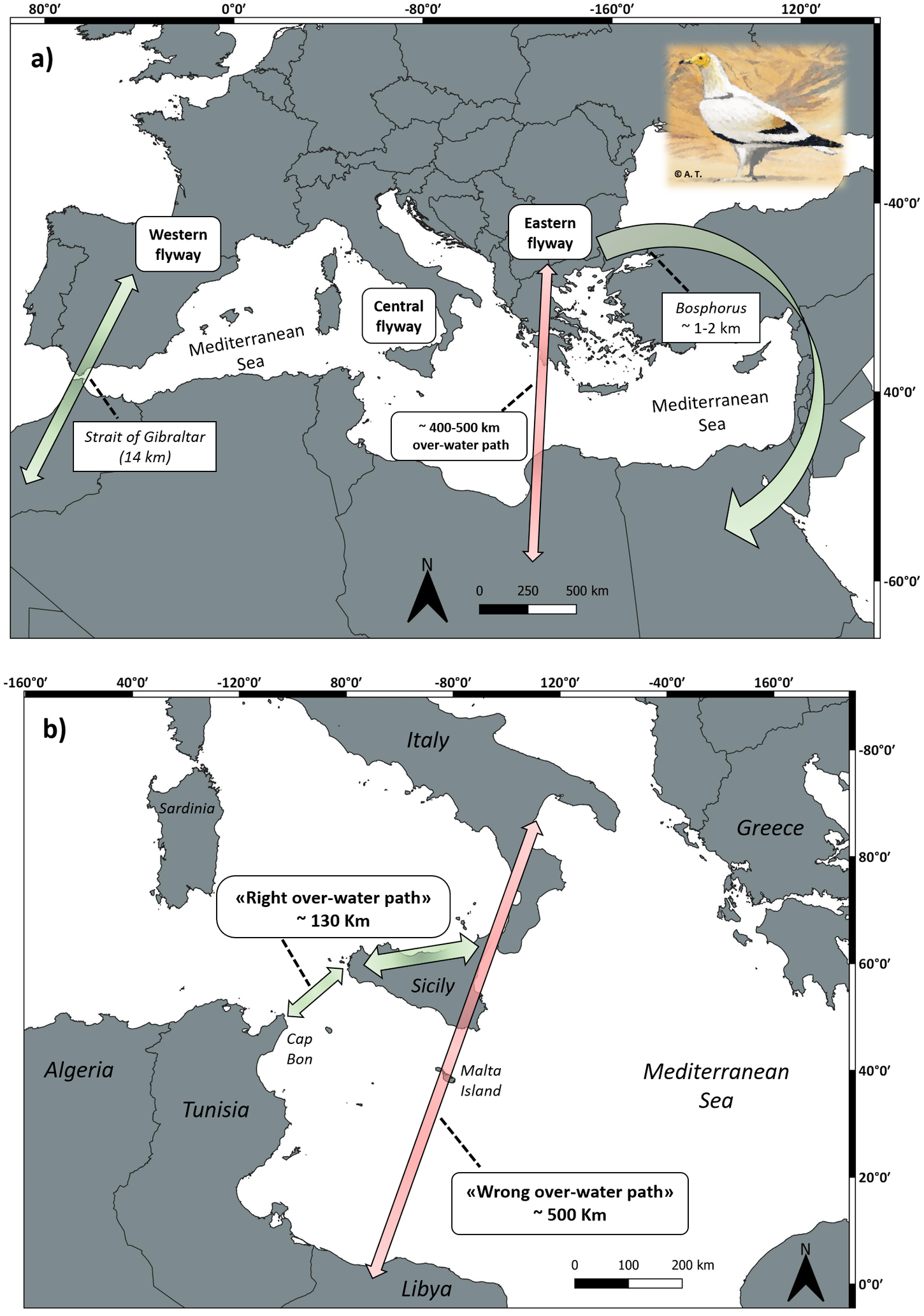
Figure 1. Schematic representation of the migratory flyways followed by Egyptian Vulture populations breeding in the Western Palearctic. Green arrows represent “right” migration paths minimizing sea-crossing, while red arrows represent “wrong” paths within a flyway including the crossing of larger sea tracts over the Mediterranean Sea. Approximate distance of such over-water paths is reported in km. Panel a) the western flyway is followed by individuals breeding in Southern France and the Iberian Peninsula across the Strait of Gibraltar and towards wintering areas in West Africa; the eastern flyway is followed by individuals breeding in the Balkans and Turkey through the eastern coast of the Mediterranean and towards the Red Sea and wintering in eastern sub-Saharan Africa. Panel b) zoom on the central flyway which is followed by individuals of the relict Italian population across Southern continental Italy and Sicily towards African coasts. The drawing of the Egyptian Vulture is by Alessandro Troisi.
In general, migratory peaks occur between mid-August and mid-September in autumn and the second half of March and early April during spring (Oppel et al. Reference Oppel, Buechley, López-López, Phipps, Arkumarev, Bounas, Williams, Dobrev, Dobrev, Stoychev, Kret, Cenerini, Ceccolini, Saravia, Nikolov, Panuccio, Mellone and Agostini2021b). Juveniles usually spend about three to four years roaming in Africa before returning to Europe for the first time, once sexual maturity is reached. Owing to its weight and morphology, the energy expenditure associated with the active flapping flight is demanding, especially over the open sea and if protracted in time (Agostini et al. Reference Agostini, Panuccio and Pasquaretta2015). As a result, birds breeding in Europe tend to migrate over land which keeps them away from the open sea: those migrating across the western flyway funnel through the Strait of Gibraltar (14–20 km wide), and those travelling across the eastern flyway through the Bosphorus (which is only 1–2 km wide, see Oppel et al. Reference Oppel, Dobrev, Arkumarev, Saravia, Bounas, Kret, Velevski, Stoychev and Nikolov2015; Fig. 1).
In studies aiming at investigating differential survival between Egyptian Vulture populations throughout the full annual cycle (Buechley et al. Reference Buechley, Oppel, Efrat, Phipps, Carbonell Alanís, Álvarez, Andreotti, Arkumarev, Berger-Tal, Bermejo Bermejo, Bounas, Ceccolini, Cenerini, Dobrev, Duriez, García, García-Ripollés, Galán, Gil, Giraud, Hatzofe, Iglesias-Lebrija, Karyakin, Kobierzycki, Kret, Loercher, López-López, Miller, Mueller, Nikolov, de la Puente, Sapir, Saravia, Sekercioglu, Sillett, Tavares, Urios and Marra2021, Oppel et al. Reference Oppel, Saravia, Bounas, Arkumarev, Kret, Dobrev, Dobrev, Kordopatis, Skartsi, Velevski, Petrovski, Bino, Topi, Klisurov, Stoychev and Nikolov2021c) it was found that: i) lower survival mostly occurred during migratory periods and; ii) there were proportionally fewer confirmed mortalities in the western flyway compared to the central and eastern one (in Buechley et al. Reference Buechley, Oppel, Efrat, Phipps, Carbonell Alanís, Álvarez, Andreotti, Arkumarev, Berger-Tal, Bermejo Bermejo, Bounas, Ceccolini, Cenerini, Dobrev, Duriez, García, García-Ripollés, Galán, Gil, Giraud, Hatzofe, Iglesias-Lebrija, Karyakin, Kobierzycki, Kret, Loercher, López-López, Miller, Mueller, Nikolov, de la Puente, Sapir, Saravia, Sekercioglu, Sillett, Tavares, Urios and Marra2021, individuals from Italy, i.e. the central flyway, were included in the “eastern” group). iii) captive-bred juveniles released in the Balkans in their second calendar year, have shown higher survival probability than those released in their first calendar year, as well as than wild-hatched juveniles (Oppel et al. Reference Oppel, Saravia, Bounas, Arkumarev, Kret, Dobrev, Dobrev, Kordopatis, Skartsi, Velevski, Petrovski, Bino, Topi, Klisurov, Stoychev and Nikolov2021c). Interestingly, unlike birds released in the Balkans, a recent study has shown no difference in apparent survival between captive-bred and wild birds in the Egyptian Vulture population in Israel (Efrat et al. Reference Efrat, Hatzofe, Miller, Mueller, Sapir and Berger-Tal2022). The differences in survival between these populations may stem from differences in migration routes and related risks, since Egyptian Vultures from Israel do not have to cope with large over-water crossing during migration (Efrat et al. Reference Efrat, Hatzofe, Miller, Mueller, Sapir and Berger-Tal2022).
Egyptian Vultures migrating along the eastern flyway cover distances that are less direct and up to twice as long as those covered by vultures from Western Europe (Phipps et al. Reference Phipps, López-López, Buechley, Oppel, Álvarez, Arkumarev, Bekmansurov, Berger-Tal, Bermejo, Bounas, Alanís, de la Puente, Dobrev, Duriez, Efrat, Fréchet, García, Galán, García-Ripollés, Gil, Iglesias-Lebrija, Jambas, Karyakin, Kobierzycki, Kret, Loercher, Monteiro, Morant Etxebarria, Nikolov, Pereira, Peške, Ponchon, Realinho, Saravia, Şekercioğlu, Skartsi, Tavares, Teodósio, Urios and Vallverdú2019). In addition, not all juveniles from the Balkan Peninsula pass through the Bosphorus: as shown by Oppel et al. (Reference Oppel, Dobrev, Arkumarev, Saravia, Bounas, Kret, Velevski, Stoychev and Nikolov2015), out of 19 birds tracked, 10 did not follow older birds heading towards the Bosphorus, but failed the migration route. Among them, only one survived (Oppel et al. Reference Oppel, Dobrev, Arkumarev, Saravia, Bounas, Kret, Velevski, Stoychev and Nikolov2015). Presumably these birds moved along an inherited N-S axis undertaking a long sea-crossing between Southern Greece and Libya (approximately 400–500 km, similar in length to that between Southern Sicily and Libya; Fig. 1b). This latter population is also experiencing marked declines and range contractions (Oppel et al. Reference Oppel, Buechley, López-López, Phipps, Arkumarev, Bounas, Williams, Dobrev, Dobrev, Stoychev, Kret, Cenerini, Ceccolini, Saravia, Nikolov, Panuccio, Mellone and Agostini2021b). For instance, a dramatic decline occurred in the Greek population, from 250 pairs in the 1970s to a minimum of only 12 pairs recorded in 2013 (Velevski et al. Reference Velevski, Nikolov, Hallmann, Dobrev, Sidiropoulos, Saravia, Tsiakiris, Arkumarev, Galanaki, Konimos, Kalliopi, Kret, Grubač, Lisičanec, Kastritis, Vavylis, Topi, Hoxha and Oppel2015), while less than 70 pairs remain in the whole Balkan Peninsula (Keller et al. Reference Keller, Herrando, Voříšek, Franch, Kipson, Milanesi, Martí, Anton, Klvaňová, Kalyakin, Bauer and Foppen2020).
In Southern Italy, due to peculiar geographical conditions, Egyptian Vultures are forced to undertake the long sea-crossing between Sicily and North Africa to reach their African wintering grounds in Central-Western Sahel (Agostini et al. Reference Agostini, Logozzo and Panuccio2000, Reference Agostini, Premuda, Mellone, Panuccio, Logozzo, Bassi and Cocchi2004, Ceccolini and Cenerini Reference Ceccolini and Cenerini2016). As expected for a typical soaring migrant, in this area passage is concentrated through the Channel of Sicily, between Western Sicily and the Cap Bon Peninsula (Tunisia), and in particular over the island of Marettimo (Agostini et al. Reference Agostini, Logozzo and Panuccio2000, Reference Agostini, Premuda, Mellone, Panuccio, Logozzo, Bassi and Cocchi2004) used as a stepping-stone before a non-stop flapping flight over the open sea of at least 130 km (Figs. 1, 2). This over-water journey is the shortest available through the central flyway (i.e. the flyway to minimize risks for a typical soaring species). Other routes over the Mediterranean Sea represent larger paths that can reach up to c.500 km (Fig. 1), which means highly risky obstacles to cope with that could be lethal for juveniles. In addition, these latter routes can also pass over Malta, a well-known hotspot for wildlife crime, where illegal shooting still takes place and affects many protected species, including Egyptian Vultures (Fenech Reference Fenech1992, www.lifegyptianvulture.it).
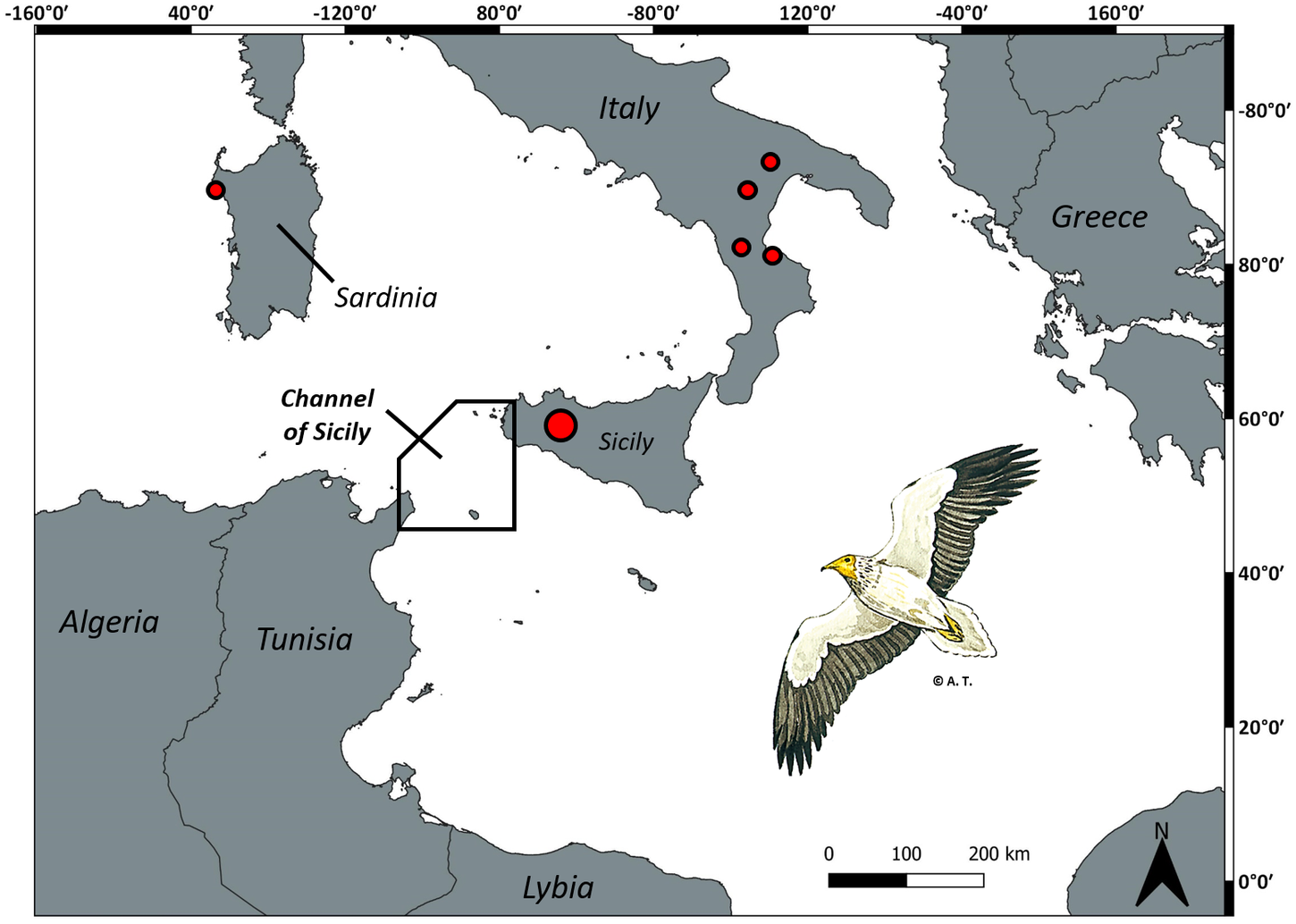
Figure 2. The central-eastern Mediterranean. The solid-line polygon over the Channel of Sicily delimits the area over which wind statistics in Fig. 4 were computed. The red dots indicate the five breeding sites in Southern Italy as well as the single pair in Sardinia. Size of red dots is proportional to the number of breeding pairs (refer to main text). The drawing of the Egyptian Vulture is by Alessandro Troisi.
Similar to Balkan populations, the Egyptian Vulture in Italy was once widely distributed across central to southern regions (including Sicily) and comprised c.50 pairs in the 1970s (Brichetti and Fracasso Reference Brichetti and Fracasso2018). Nowadays, the species is considered critically endangered on the national red list (Gustin et al. Reference Gustin, Nardelli, Brichetti, Battistoni, Rondinini and Teofili2019). In the last few decades, the breeding population has dropped to less than 10–12 breeding pairs (Keller et al. Reference Keller, Herrando, Voříšek, Franch, Kipson, Milanesi, Martí, Anton, Klvaňová, Kalyakin, Bauer and Foppen2020) mainly due to poaching, disturbance of nesting sites, illegal poisoning, and reduction in their food supply, and it is now exposed to the causal factors that lead to the “extinction vortex” (Andreotti and Leonardi Reference Andreotti and Leonardi2009). The last breeding pairs are patchily distributed across four regions in the south: 3–4 pairs between Apulia, Basilicata and Calabria regions and another 7–8 pairs concentrated in Sicily (Fig. 2; Brichetti and Fracasso Reference Brichetti and Fracasso2018). It should be noted that, since 2019, a single pair settled in Sardinia (Fig. 2), in Central Italy, far away from those in the south.
The case of restocking Egyptian Vulture in Italy
To promote the demographic recovery and expansion of the critically endangered Italian Egyptian Vulture population, a captive-breeding programme started in 1993, with the first releases in 2003 (Andreotti and Leonardi Reference Andreotti and Leonardi2009, Ceccolini and Cenerini Reference Ceccolini and Cenerini2016, Progetto Capovaccaio Reference Capovaccaio2022). Most of the birds used were captive-born in the breeding facilities of the CERM association (Endangered Raptors Centre – Rocchette di Fazio, GR, Tuscany, Italy), while some others originated from European zoos, such as those of Jerez de La Frontera (Spain), Prague, and Zlin (Czech Republic) (Ceccolini and Cenerini Reference Ceccolini and Cenerini2016). Since 2017, the programme has benefitted from the support of an EU project: “Life Egyptian Vulture - Measures for the conservation of the Egyptian Vulture in Italy and the Canary Islands – LIFE16 NAT/IT/000659 (www.lifegyptianvulture.it). The ongoing project envisages a series of concrete actions to mitigate the impact of negative factors affecting the species, such as: the surveillance of nesting sites, the securing of power lines for the mitigation of electrocution, activities to combat the illegal use of poison, the construction and management of supplementary feeding stations, as well as the management of ex-situ reproduction and release of captive-bred individuals.
Overall, three different release techniques were adopted by the programme: fostering (used only in one case in 2003; Ceccolini and Cenerini Reference Ceccolini and Cenerini2005, Di Vittorio et al. Reference Di Vittorio, Falcone, Diliberto, Cortone, Massa, Sarà and Sergio2006), hacking, and delayed-release (Ceccolini and Cenerini Reference Ceccolini and Cenerini2005, Progetto Capovaccaio Reference Capovaccaio2022). The majority of birds were released using the hacking technique: birds were put into the hacking facilities and then released into the wild in Southern Italy, where the last wild breeding pairs are located (Fig. 2). In particular, release operations started to (and will) be carried out using two alternative methods: i) release during the summer (second half of August) of fledglings born in captivity during the spring (thus at an approximate age of 3–4 months), after a brief period of settling in the release area inside aviaries or caged natural cavities; ii) release during the spring of juveniles born in captivity during the previous year (therefore over the age of one) after a few weeks settling in the release area inside the aviaries.
According to official published and public data, almost 40 captive-bred individuals were released, as part of the restocking programme conducted between 2003 and 2021 (Ceccolini and Cenerini Reference Ceccolini and Cenerini2005, Reference Ceccolini and Cenerini2016, Ceccolini et al. Reference Ceccolini, Cenerini and Aebischer2009, Progetto Capovaccaio Reference Capovaccaio2022, www.lifegyptianvulture.it). All but three (one released in Sicily in 2003 and two in Tuscany in 2005; Ceccolini and Cenerini Reference Ceccolini and Cenerini2005, Andreotti and Leonardi Reference Andreotti and Leonardi2009, Progetto Capovaccaio Reference Capovaccaio2022) were released in Southern continental Italy (Fig. 2). Among these birds, 21 provided information about their first migratory pathway through the Central Mediterranean, after being equipped with satellite transmitters or being re-observed (Ceccolini and Cenerini Reference Ceccolini and Cenerini2005, Reference Ceccolini and Cenerini2016, Andreotti and Leonardi Reference Andreotti and Leonardi2009, Ceccolini et al. Reference Ceccolini, Cenerini and Aebischer2009, Brichetti and Fracasso Reference Brichetti and Fracasso2018, Progetto Capovaccaio Reference Capovaccaio2022, www.lifegyptianvulture.it). We did not consider an individual that, after having been released at the age of one year and having spent the first two winters in Southern Italy, started wandering around the Balkans and then embarked on its first autumn migration at the age of three via another wrong pathway, that of the eastern flyway (between Greece and Libya, Fig. 1a), managing to reach the African coast (www.lifegyptianvulture.it).
Among these 21 birds, 12 (57.1%) took the wrong pathway. Once arrived in Eastern Sicily, these birds (mostly 1st calendar year individuals) moved southwards and crossed the Mediterranean Sea along the longer path (between south-eastern Sicily and Libya) that led them to fly over 500 km over the sea (Fig. 1b). Six out of these 12 (50%) survived the long crossing and reached Africa. One bird (8.33%) disappeared over Malta (probably killed) and the other five (41.66%) fell into the sea and died.
Conversely, among the remaining nine juveniles (42.9%) who used the right pathway, all were successful (88.9%) except one (11.1%). While the latter died in the sea, the former crossed the Channel of Sicily successfully and reached Tunisia (Ceccolini and Cenerini Reference Ceccolini and Cenerini2016, Progetto Capovaccaio Reference Capovaccaio2022, www.lifegyptianvulture.it).
Such data, together with those concerning juveniles born in the Balkan Peninsula and crossing between Southern Greece and Libya, clearly show how paths involving hundreds of kilometres over the Mediterranean Sea represent one of the most prominent natural barriers reducing survival: every year many juveniles drown in the Mediterranean on their first autumn migration, and this mortality is higher than that recorded in the Sahara Desert (e.g. Buechley et al. Reference Buechley, Oppel, Efrat, Phipps, Carbonell Alanís, Álvarez, Andreotti, Arkumarev, Berger-Tal, Bermejo Bermejo, Bounas, Ceccolini, Cenerini, Dobrev, Duriez, García, García-Ripollés, Galán, Gil, Giraud, Hatzofe, Iglesias-Lebrija, Karyakin, Kobierzycki, Kret, Loercher, López-López, Miller, Mueller, Nikolov, de la Puente, Sapir, Saravia, Sekercioglu, Sillett, Tavares, Urios and Marra2021). Conversely, where Egyptian Vultures do not have to cross such large over-water paths (e.g. Israel), apparent survival is not altered by the release age or season (Efrat et al. Reference Efrat, Hatzofe, Miller, Mueller, Sapir and Berger-Tal2022).
The importance of wind patterns
The central part of the Mediterranean Sea represents a difficult barrier to this typical soaring species also at its shorter point because of regional wind regimes. Indeed, wind conditions between the Cap Bon Peninsula in Tunisia and Western Sicily appear often suboptimal for water-crossing both in early spring (second half of March) and late summer (mid-August to mid-September), when migration peaks of the species in the Mediterranean area occur (Finlayson Reference Finlayson1992, Oppel et al. Reference Oppel, Buechley, López-López, Phipps, Arkumarev, Bounas, Williams, Dobrev, Dobrev, Stoychev, Kret, Cenerini, Ceccolini, Saravia, Nikolov, Panuccio, Mellone and Agostini2021b). In particular, we analysed wind data in the period 1991–2020 obtained from the ERA5 reanalysis product provided by the Copernicus Climate Change Service (C3S) (Hersbach et al. 2018), providing a reconstruction of historical atmospheric conditions at 0.25° resolution. Hourly winds on pressure levels were averaged in the range 1,000–900 hPa, corresponding to about the first 1,000 m in height, encompassing the range of altitudes covered by migrating raptors over water (Kerlinger Reference Kerlinger1989). All wind statistics were computed between 08h00 and 16h00, the time range in which most large soaring raptors actively migrate (Mellone et al. Reference Mellone, Klaassen, Garcia-Ripolles, Limiñana, López-López and Pavón2012). Although winds with variable directions blow over the Channel during both spring and autumn, favourable (tail) winds (coming from SW in spring and from NE in autumn) rarely occur. In particular, winds are stronger (average modulus of hourly wind: 8.64 m/s) and with a higher frequency of north-westerly winds in spring, during the second half of March. In this period, they can frequently exceed 10 m/s (Figs. 3a, 4a). On the other hand, in autumn winds are weaker and seldom exceed 10 m/s, with a higher frequency of southerly winds (headwinds), during both the second half of August (average modulus of hourly wind: 5.05 m/s) and the first half of September (5.88 m/s; Figs. 3c, d, 4c, d).

Figure 3. Wind patterns in the Channel of Sicily in late March (a), early April (b), late August (c) and early September (d) in the years from 1991 to 2020. The colour map indicates the average value of the modulus of the hourly wind; the arrows represent the composite hourly wind vector over the averaging period. The letter “M” labels the island of Marettimo. Wind data were obtained from the ERA5 reanalysis product, as described in the text.

Figure 4. Joint probability density function (pdf) of wind origin angle and wind modulus for hourly wind in the Channel of Sicily in late March (a), early April (b), late August (c) and early September (d) in the years from 1991 to 2020. The abscissa represents the wind origin direction, for example ‘NW’ indicates a north-westerly wind. The colour map represents the probability density of the joint pdf. The area considered for deriving this statistic is delimited with a black polygon in Figure 2. Wind data were obtained from the ERA5 reanalysis product, as described in the text.
Interestingly, Egyptian Vultures facing the Strait of Gibraltar, the short water-crossing between Spain and Morocco, do not cross during strong lateral winds, which limits the risk of dying over the open water, especially when drifted towards the Atlantic Ocean by easterly winds (Finlayson Reference Finlayson1992). If Egyptian Vultures hesitate at this relatively short water-crossing and wait for better wind conditions, a more conservative strategy is expected in birds facing the 10-fold longer water-crossing through the Channel of Sicily, where the risk of drowning is potentially higher and wind patterns are often suboptimal for crossing. In addition, the longer the crossing, the higher the risk of unpredictable weather changes during the flight over water. The high frequency of suboptimal wind conditions over the Channel of Sicily could be detrimental to breeders especially in early spring. As a matter of fact, if protracted in time, adverse weather conditions over the Channel might delay the beginning of the breeding season and consequently the beginning of the post-fledging migration of juveniles from these wild pairs. In these circumstances, a consequent detrimental effect on the social learning of juveniles can occur. In fact, juveniles would have less time to improve flight skills, while missing their parents that start migrating few weeks after fledging (Oppel et al. Reference Oppel, Buechley, López-López, Phipps, Arkumarev, Bounas, Williams, Dobrev, Dobrev, Stoychev, Kret, Cenerini, Ceccolini, Saravia, Nikolov, Panuccio, Mellone and Agostini2021b). Since the parents are expected to migrate without their offspring, young Egyptian Vultures cannot follow them on their first migration (Oppel et al. Reference Oppel, Buechley, López-López, Phipps, Arkumarev, Bounas, Williams, Dobrev, Dobrev, Stoychev, Kret, Cenerini, Ceccolini, Saravia, Nikolov, Panuccio, Mellone and Agostini2021b). Thus, if adverse wind conditions in early spring delay the beginning of the breeding season (and juveniles’ post-fledging migration), we suspect that chances of meeting older birds en route are unlikely. Frequent suboptimal wind conditions over the Channel of Sicily occur during the second half of March persisting into the first half of April (Figs. 3b, 4b). Egg-laying by the Egyptian Vulture usually occurs in early April (Abuladze and Shergalin Reference Abuladze, Shergalin, Chancellor, Meyburg and Ferrero1998, Gensbol Reference Gensbøl2008), but in Southern continental Italy it has been reported later in the season (median date 23 April; Brichetti and Fracasso Reference Brichetti and Fracasso2018). Finally, since birds returning north and heading towards the Channel of Sicily have to fight the prevailing north-easterly winds over the Sahara Desert (Mellone et al. Reference Mellone, Klaassen, Garcia-Ripolles, Limiñana, López-López and Pavón2012, Panuccio et al. Reference Panuccio, Mellone and Agostini2021), they are expected to stop migrating for some days to refuel along north African coasts (as shown by Buechley et al. Reference Buechley, Oppel, Efrat, Phipps, Carbonell Alanís, Álvarez, Andreotti, Arkumarev, Berger-Tal, Bermejo Bermejo, Bounas, Ceccolini, Cenerini, Dobrev, Duriez, García, García-Ripollés, Galán, Gil, Giraud, Hatzofe, Iglesias-Lebrija, Karyakin, Kobierzycki, Kret, Loercher, López-López, Miller, Mueller, Nikolov, de la Puente, Sapir, Saravia, Sekercioglu, Sillett, Tavares, Urios and Marra2021) before the long sea crossing, further delaying the start of the breeding season.
The role of guidance and meeting opportunities
Traditional migration systems where juveniles reduce the difficulty of navigation by following experienced birds may no longer function in low-density or declining populations as well as in large and well-structured populations (Oppel et al. Reference Oppel, Buechley, López-López, Phipps, Arkumarev, Bounas, Williams, Dobrev, Dobrev, Stoychev, Kret, Cenerini, Ceccolini, Saravia, Nikolov, Panuccio, Mellone and Agostini2021b). In addition, natal dispersal distances are known to be inversely related to vulture density in the natal populations, suggesting that birds perceive the abundance of conspecifics as a signal of habitat quality (Serrano et al. Reference Serrano, Cortés-Avizanda, Zuberogoitia, Blanco, Benítez, Ponchon, Grande, Ceballos, Morant, Arrondo, Zabla, Montelío, Ávila, González, Arroyo, Frías, Kobierzycki, Arenas, Tella and Donázar2021). This is particularly alarming for low-density and declining populations, such as the case of the Italian population, where a substantial number of juvenile birds (either wild-born or captive-bred) may struggle to find the right flyway and associate with experienced migrants because of the numerical scarcity and their patchy distribution (Fig. 2; Andreotti and Leonardi Reference Andreotti and Leonardi2009).
Indeed, during observations made over Marettimo Island between the end of August and mid-September in 1997, 1998 and 2003, nine (5.5%, n = 163) juvenile Egyptian Vultures were counted, compared to 141 (86.5%) adults and 13 (8%) sub-adults (Agostini et al. Reference Agostini, Logozzo and Panuccio2000, Reference Agostini, Premuda, Mellone, Panuccio, Logozzo, Bassi and Cocchi2004). At that time, the Italian population was estimated at 9–13 breeding pairs, and at least 19–22 juveniles were expected to migrate in the three years (Liberatori and Penteriani Reference Liberatori and Penteriani2001, Di Vittorio et al. Reference Di Vittorio, Diliberto and Campobello2003, Sarà et al. Reference Sarà, Grenci and Di Vittorio2009). Surveys made at Marettimo reported that all juveniles were migrating in flock with adults, and sometimes in mixed-species flocks with adult European Honey Buzzards Pernis apivorus and Black Kites Milvus migrans (Agostini et al. Reference Agostini, Logozzo and Panuccio2000, Reference Agostini, Premuda, Mellone, Panuccio, Logozzo, Bassi and Cocchi2004). In fact, every year, hundreds of Honey Buzzards and Black Kites migrate in flocks along this route from Western Sicily to Cap Bon (Agostini et al. Reference Agostini, Logozzo and Panuccio2000), thus representing a good chance for inexperienced juvenile Egyptian Vultures to find the right path by following them. In contrast, on the wrong path (Fig. 1b), the risk that juveniles accidentally fail by venturing over 500 km of open sea is high because of the impossibility of joining a flock. According to data shown by Ceccolini and Cenerini (Reference Ceccolini and Cenerini2016), the same could occur with captive-bred young vultures released in late summer in Southern continental Italy.
Conversely, in the case of flyways not including a large sea-crossing, mortality rates are lower (Buechley et al. Reference Buechley, Oppel, Efrat, Phipps, Carbonell Alanís, Álvarez, Andreotti, Arkumarev, Berger-Tal, Bermejo Bermejo, Bounas, Ceccolini, Cenerini, Dobrev, Duriez, García, García-Ripollés, Galán, Gil, Giraud, Hatzofe, Iglesias-Lebrija, Karyakin, Kobierzycki, Kret, Loercher, López-López, Miller, Mueller, Nikolov, de la Puente, Sapir, Saravia, Sekercioglu, Sillett, Tavares, Urios and Marra2021). For example, juveniles from Spain and France have lower mortality rates during migration because their birth places are located along a safer flyway that funnels them towards the Strait of Gibraltar even when they do not follow experienced individuals (Phipps et al. Reference Phipps, López-López, Buechley, Oppel, Álvarez, Arkumarev, Bekmansurov, Berger-Tal, Bermejo, Bounas, Alanís, de la Puente, Dobrev, Duriez, Efrat, Fréchet, García, Galán, García-Ripollés, Gil, Iglesias-Lebrija, Jambas, Karyakin, Kobierzycki, Kret, Loercher, Monteiro, Morant Etxebarria, Nikolov, Pereira, Peške, Ponchon, Realinho, Saravia, Şekercioğlu, Skartsi, Tavares, Teodósio, Urios and Vallverdú2019, Buechley et al. Reference Buechley, Oppel, Efrat, Phipps, Carbonell Alanís, Álvarez, Andreotti, Arkumarev, Berger-Tal, Bermejo Bermejo, Bounas, Ceccolini, Cenerini, Dobrev, Duriez, García, García-Ripollés, Galán, Gil, Giraud, Hatzofe, Iglesias-Lebrija, Karyakin, Kobierzycki, Kret, Loercher, López-López, Miller, Mueller, Nikolov, de la Puente, Sapir, Saravia, Sekercioglu, Sillett, Tavares, Urios and Marra2021). In agreement with this assumption, among aged birds at this bottleneck, 64% were adults (vs 86.5% over Marettimo) and 27.3% juveniles (vs 5.5% over Marettimo), resulting in 0.9 juveniles per pair (Finlayson Reference Finlayson1992) which is even higher than the breeding success of the species (Ferguson-Lees and Christie Reference Ferguson-Lees and Christie2001).
It is possible that low population density, and potential delay in the breeding season due to adverse wind patterns, may have further indirectly exacerbated the decline in the Italian population, mainly due to anthropogenic threats such as electrocution, illegal shooting, and poisoning (Sarà and Di Vittorio Reference Sarà and Di Vittorio2003, Agostini et al. Reference Agostini, Panuccio and Pasquaretta2015). In this scenario, conspecific meeting opportunities and social learning appear even more difficult for the captive-bred juvenile Egyptian Vultures released in Southern continental Italy, especially if released late in the season. Considering that egg laying has been recorded in late April for pairs breeding in Southern continental Italy (Brichetti and Fracasso Reference Brichetti and Fracasso2018), several wild juveniles born in early June (incubation period lasts about 42 days; Ferguson-Lees and Christie Reference Ferguson-Lees and Christie2001) are expected to leave their natal site late in the season, thus decreasing even more their chances to meet older birds that have already left at that time. This would further explain the relatively low number of juveniles (about the half of those expected) observed in flock with adults through the Channel of Sicily during post-breeding migration (Agostini et al. Reference Agostini, Logozzo and Panuccio2000, Reference Agostini, Premuda, Mellone, Panuccio, Logozzo, Bassi and Cocchi2004).
Management and conservation prospects
The Egyptian Vulture is the most threatened bird of prey in Italy, suffering from a multitude of both anthropogenic (e.g. electrocution, illegal shooting, illegal poison, direct disturbance) and natural threats and with a still declining population. To halt this trend and boost population persistence over time, specific conservation measures are required together with proactive management actions. As unfavourable winds usually blow during migration periods (especially in spring), both the location and date of release of captive-bred Egyptian Vultures should be carefully evaluated to fit with increasing chances of a) merging with other migrating birds enabling them to choose the shortest over-water path and b) promoting philopatry towards higher population density areas.
To further limit the chance that captive-bred released birds use the wrong pathway (between Southern Sicily and Libya), it would probably be helpful to release individuals (especially if inexperienced 1st calendar birds) in Central-Western Sicily, rather than in Southern continental Italy (Apulia, Calabria or Basilicata regions). Therefore, released juveniles may have even more chances to find and select the safer route by meeting adults belonging to the local breeding population (almost 70% of Italian breeding pairs reside in Sicily) or adults of other species, such as European Honey Buzzards and Black Kites, passing in large flocks through Western Sicily en route to the Channel of Sicily between mid-August and early September. Furthermore, because of the species’ philopatry, released juveniles would tend, once sexually mature, to return to Sicilian release sites where the higher vulture population density could potentially serve as a signal of habitat quality (and social information could help individuals to select where to settle; Serrano et al. Reference Serrano, Cortés-Avizanda, Zuberogoitia, Blanco, Benítez, Ponchon, Grande, Ceballos, Morant, Arrondo, Zabla, Montelío, Ávila, González, Arroyo, Frías, Kobierzycki, Arenas, Tella and Donázar2021).
It is also recommended to release captive-bred individuals earlier in the season, rather than at the end of it in late August, by using 2st calendar year birds. In the case of particular technical circumstances that still require releases in Southern continental Italy (Apulia, Basilicata and Calabria), it is still recommended to release 2st year birds as a priority as they are expected to have higher survival probability (Oppel et al. Reference Oppel, Saravia, Bounas, Arkumarev, Kret, Dobrev, Dobrev, Kordopatis, Skartsi, Velevski, Petrovski, Bino, Topi, Klisurov, Stoychev and Nikolov2021c) and to migrate earlier in the season (compared to fledging young), thus increasing their chances of meeting older birds. Indeed, using delayed release, captive-bred individuals can be an effective approach for successful restocking programmes (e.g. Oppel et al. Reference Oppel, Saravia, Bounas, Arkumarev, Kret, Dobrev, Dobrev, Kordopatis, Skartsi, Velevski, Petrovski, Bino, Topi, Klisurov, Stoychev and Nikolov2021c, Efrat et al. Reference Efrat, Hatzofe, Miller, Mueller, Sapir and Berger-Tal2022). Interestingly, in a recent study, Oppel et al. (Reference Oppel, Saravia, Bounas, Arkumarev, Kret, Dobrev, Dobrev, Kordopatis, Skartsi, Velevski, Petrovski, Bino, Topi, Klisurov, Stoychev and Nikolov2021c) evaluated realistic reinforcement and survival improvement scenarios, by using an integrated population model to estimate how many captive-bred Egyptian Vultures would need to be released to stabilise the Balkan population. The authors concluded that the release of 12 or more birds, preferably at least in their 2st calendar year, every year for 30 years would reduce the probability of extinction from 48% without reinforcement to <1%.
How to further limit the probability of extinction of the Italian Egyptian Vulture population in the next decades? First, by concentrating efforts on minimising the anthropogenic threats that undermine the survival of this population. In this sense, the “Life Egyptian Vulture” project is at the forefront of activating a series of direct actions to mitigate the impact of threats, ultimately aimed at halting the decline of the population (www.lifegyptianvulture.it). At the same time, it is important to continue with the release of captive-bred individuals, but primarily in areas that minimize the chance of following a wrong migration route (e.g. Central-Western Sicily rather than Southern continental Italy) and by using 2st calendar year individuals rather than inexperienced 1st year birds, in order to increase their chances of survival. The activation of release sites in Sicily seems therefore particularly urgent and of high priority, as indicated in the National Action Plan for the Egyptian Vulture (Andreotti and Leonardi Reference Andreotti and Leonardi2009), in particular for releasing young individuals who fledge later in the season and which, if released in Southern continental Italy, risk missing the train of migrants.
However, the key factor needed to proceed in this sense is the presence of: 1) a protected area with suitable habitats and potential nesting sites for the Egyptian Vulture. The protected area is essential to ensure adequate protection to the site and to the individuals before and after release, while the presence of suitable habitat characterized by low undisturbed cliffs as potential nesting site, distant from urban areas and surrounded by arboreal crops and Mediterranean vegetation (Sarà and Di Vittorio Reference Sarà and Di Vittorio2003), is important for reasons related to philopatry and site-fidelity; 2) one or more nesting pairs in the area or in its immediate vicinity, for reasons related to social information and guidance opportunities; 3) the technical availability of the protected area management to implement all the measures necessary for a successful outcome of the release operations, subsequent monitoring of the released birds, and management of the feeding stations.
The implementation of supplementary feeding stations (sensu Duriez et al. Reference Duriez, Andevski, Bowden, Camiña-Cardenal, Frey, Genero, Hatzofe, Llopis-Dell, Néouze, Phipps and Tavares2021) could increase the probability of conspecific meeting between juveniles and older birds. However, although vulture feeding stations are one of the most used and common tools for vulture conservation (Moreno-Opo et al. Reference Moreno-Opo, Trujillano and Arredondo2015, Duriez et al. Reference Duriez, Andevski, Bowden, Camiña-Cardenal, Frey, Genero, Hatzofe, Llopis-Dell, Néouze, Phipps and Tavares2021), they can also lead to a wide array of potential negative effects. It has been demonstrated that if not well-planned in space and time, the provision of predictable anthropogenic food, may drive changes in population dynamics, increasing competition with other species or promoting differential use of the provided resource according to specific populations (García-Heras et al. Reference García-Heras, Cortés-Avizanda and Donázar2013, Cortés-Avizanda et al. Reference Cortés-Avizanda, Blanco, DeVault, Markandya, Virani, Brandt and Donázar2016, Moreno-Opo et al. Reference Moreno-Opo, Trujillano and Margalida2020), as well as favouring resident behaviour. For instance, in recent years, increasing numbers of Egyptian Vultures are no longer migrating, remaining in Spain over winter (Morant et al. Reference Morant, Abad-Gómez, Álvarez, Sánchez, Zuberogoitia and López-López2020). There, they congregate at specific roosting and feeding sites throughout the winter period. The provision of predictable food sources as supplementary feeding stations seems to drive the apparent increase in the non-migratory population of Egyptian Vultures in Spain (Morant et al. Reference Morant, Abad-Gómez, Álvarez, Sánchez, Zuberogoitia and López-López2020). In this context, we cannot exclude that supplementary feeding stations might have a double effect: on the one hand, to increase the conspecific meeting opportunities, but on the other, to attract individuals for too long, risking them missing the "train" of the Honey Buzzards’ passage until the first week of September. Furthermore, although less likely in the short term, it could induce substantial numbers of Egyptian Vultures to change their migratory behaviour, finally turning into sedentary and wintering in Southern Italy, with potential side effects at population level in the medium-long term. Thus, this practice should be carefully evaluated both in space (placement of supplementary feeding stations) and time (when to activate them and how long to keep them active).
The conservation of the Italian Egyptian Vulture population requires joint efforts and coordinated actions. Despite promising steps made so far to limit anthropogenic threats, more targeted conservation measures that also take into account geographical and environmental factors are needed to enhance the restocking program in Italy towards a more goal-oriented and possibly long-lasting conservation outcome. The field of movement ecology has dramatically grown in the last years and the newly acquired knowledge on species movements is the key to improving the effectiveness of management strategies.
Acknowledgements
Our work benefited greatly from helpful discussions with Ugo Mellone and Giampiero Sammuri. We also thank the associate editor, Antoni Margalida, and two anonymous referees for their helpful comments on the manuscript.




