



Binge eating disorder (BED) is a pernicious eating disorder of unknown etiology, which is characterized by (i) frequent consumption of an objectively large amount of food in a discrete time period, (ii) a self-reported loss of control, and (iii) an absence of compensatory behaviors (American Psychiatric Association, 2013). Alongside these cardinal features, BED portends broad medical and psychiatric sequalae, including obesity (Mitchell, Reference Mitchell2016), metabolic syndrome (Hudson et al., Reference Hudson, Lalonde, Coit, Tsuang, EcElroy, Crow and Pope2010; Tanofsky-Kraff et al., Reference Tanofsky-Kraff, Shomaker, Stern, Miller, Sebring, Dellavalle and Yanovski2012), dyslipidemia, compromised cardiac function (American Psychiatric Association, 2013), and elevated suicidality (Olguin et al., Reference Olguin, Fuentes, Gabler, Guerdjikova, Keck and McElroy2017; Udo, Bitley, & Grilo, Reference Udo, Bitley and Grilo2019). Importantly, while approximately half of those receiving specialized treatments demonstrate symptom remission (Grilo & Masheb, Reference Grilo and Masheb2005a, Reference Grilo and Masheb2005b; Linardon, Reference Linardon2018), the need to identify brain-based abnormalities which underpin and maintain BED symptomatology is critical to advancing novel treatment initiatives.
Mechanistically, BED is theoretically predicated on an increased reward value around food, therefore facilitating a higher drive to eat, and diminished inhibitory control, which limits the extent to which food consummatory drives are tempered (Grilo & Masheb, Reference Grilo and Masheb2000). In support of this, clinical data reveal that hedonic eating – the intense pleasure derived from eating palatable food (Davis et al., Reference Davis, Levitan, Reid, Carter, Kaplan, Patte and Kennedy2012) – is reliably elevated among those with BED (Davis et al., Reference Davis, Levitan, Reid, Carter, Kaplan, Patte and Kennedy2012) and predicts the frequency of binge episodes (Lowe, Arigo, & Butryn, Reference Lowe, Arigo and Butryn2016; Witt & Lowe, Reference Witt and Lowe2014). Similarly, elevated food-related impulsivity (Giel, Teufel, Junne, Zipfel, & Schag, Reference Giel, Teufel, Junne, Zipfel and Schag2017) and difficulty diverting attention away from food-related cues is evident among those with BED (Schag et al., Reference Schag, Teufel, Junne, Preissl, Hautzinger, Zipfel and Giel2013b), and is thought to underpin the inability to abstain from food consumption observed in BED.
In elucidating the neurobiological correlates of the putative mechanisms underpinning BED, imaging studies have corroborated reward circuit hyper-responsivity during food cue presentation (Lee, Namkoong, & Jung, Reference Lee, Namkoong and Jung2017; Weygandt, Schaefer, Schienle, & Haynes, Reference Weygandt, Schaefer, Schienle and Haynes2012), which accords with elevated subjective reward sensitivity (Appelhans et al., Reference Appelhans, Woolf, Pagoto, Schneider, Whited and Liebman2011; Schienle, Schäfer, Hermann, & Vaitl, Reference Schienle, Schäfer, Hermann and Vaitl2009) and symptom severity (Wang et al., Reference Wang, Geliebter, Volkow, Telang, Logan, Jayne and Fowler2011). In concert, tasks of inhibitory control are characterized by hypoactivity in regions implicated in the inhibitory control network among those with BED, such as the dorsolateral prefrontal cortex (dlPFC) and inferior frontal gyrus (IFG) (Balodis et al., Reference Balodis, Molina, Kober, Worhunsky, White, Sinha and Potenza2013). Moreover, resting state activity among those with BED suggests a diffuse functional dysconnectivity between nodes of reward and inhibitory controls circuits in both adults and children (Haynos et al., Reference Haynos, Camchong, Pearson, Lavender, Mueller, Peterson and Lim2021; Murray et al., Reference Murray, Alba, Duval, Nagata, Cabeen, Lee and Jann2022a). Studies assessing cortical and subcortical structural characteristics of BED are relatively sparse. A recent study of children with BED revealed diffusely elevated gray matter density (GMD) in prefrontal, parietal, and temporal regions, relative to matched control children (Murray et al., Reference Murray, Duval, Balkchyan, Cabeen, Nagata, Toga and Jann2022b). With specific regards to the dlPFC, this region has been the principal site of targeted neuromodulation studies for BED, including both transcranial magnetic stimulation and transcranial direct current stimulation (Dalton, Bartholdy, Campbell, & Schmidt, Reference Dalton, Bartholdy, Campbell and Schmidt2018). Preliminary evidence suggests ameliorated BED symptom severity as a result of targeted dlPFC stimulation (Dalton et al., Reference Dalton, Bartholdy, Campbell and Schmidt2018), with one study noting altered response inhibition in those with BED (Max, Plewnia, Zipfel, Giel, & Schag, Reference Max, Plewnia, Zipfel, Giel and Schag2020). Importantly, however, almost all studies of TMS in BED have assessed exclusively female samples of adults. Further exploration of the relationship between dlPFC GMD and the putative mechanisms underpinning BED (i.e. altered behavioral approach and avoidance) may provide further insights into emerging neuromodulation treatment paradigms in both males and females, respectively.
Importantly, no studies to date have assessed sex differences in the neurobiology of BED. Historically, eating disorders in males have been assumed to represent only a small minority of all cases, and their proposed atypicality has led to their exclusion from almost all neuroimaging studies of EDs to date (Murray et al., Reference Murray, Nagata, Griffiths, Calzo, Brown, Mitchison and Mond2017). As a result, the majority of our understanding of the neurobiology of EDs stems from almost entirely female samples, and no studies to date have directly assessed sex differences in brain structure or function among those with EDs. Addressing these gaps will inform tailored treatments for males (Ganson, Murray, & Nagata, Reference Ganson, Murray and Nagata2021). In the context of BED, up to 43% of adults with BED are males (Hay, Girosi, & Mond, Reference Hay, Girosi and Mond2015), whereas in children, up to 57% of all cases are represented by males (Murray, Ganson, Chu, Jann, & Nagata, Reference Murray, Ganson, Chu, Jann and Nagata2022c). Importantly, emerging evidence has suggested nuanced differences in the clinical presentation and maintaining mechanisms of BED across males and females, respectively. For instance, and of critical importance to diagnostic criteria, males and female typically differ in conceptualizing what constitutes a binge episode. Males typically report a greater volume of food when reporting what represents a binge episode (Murray et al., Reference Murray, Nagata, Griffiths, Calzo, Brown, Mitchison and Mond2017), whereas females are more likely to report a loss of control during binge episodes (Reslan & Saules, Reference Reslan and Saules2011). Moreover, females with BED more commonly associate binge episodes with negative affect (Reslan & Saules, Reference Reslan and Saules2011).
Cumulatively, these data raise the intriguing possibility of sex differences in the neurobiological correlates of BED. This is especially salient to examine in pre-adolescent and adolescent populations, where the prevalence of BED gradually shifts from a relatively even distribution between sexes, to a post-pubertal skew in prevalence towards females (Mikhail et al., Reference Mikhail, Anaya, Culbert, Sisk, Johnson and Klump2021). To that end, assessing GMD may be an important starting point in this line of inquiry, given (i) the differential rates of gray matter development in boys and girls throughout adolescence, respectively (Gennatas et al., Reference Gennatas, Avants, Wolf, Satterthwaite, Ruparel, Ciric and Gur2017), and (ii) evidence noting perturbations in GMD among children with BED, relative to controls (Murray et al., Reference Murray, Alba, Duval, Nagata, Cabeen, Lee and Jann2022a, Reference Murray, Duval, Balkchyan, Cabeen, Nagata, Toga and Jann2022b, Reference Murray, Ganson, Chu, Jann and Nagata2022c). However, given that gray matter morphometric changes evolve rapidly throughout adolescence, and in differential trajectories among boys and girls (Gennatas et al., Reference Gennatas, Avants, Wolf, Satterthwaite, Ruparel, Ciric and Gur2017), the need to map neurodevelopmental changes throughout adolescence ought to be underscored. Notwithstanding, assessing potential sex differences in GMD in BED at an early age, and delineating how potential differences relate to putative mechanisms involves in BED psychopathology, may share important insights into emerging treatment development. For instance, the broader assessment of sex differences in neurobiological mechanisms underpinning BED may provide important insights around how neuromodulation treatments are developed and disseminated among boys and girls, respectively.
The present study therefore aimed to elucidate sex differences in cortical and subcortical gray matter characteristics associated with BED among pre-adolescent boys and girls, where disease prevalence is most comparably distributed across genders. In addition, we aimed to examine the relationship between one region of brain which reliably demonstrates structural (Murray et al., Reference Murray, Duval, Balkchyan, Cabeen, Nagata, Toga and Jann2022b) and functional (Murray et al., Reference Murray, Alba, Duval, Nagata, Cabeen, Lee and Jann2022a) abnormalities in children with BED - the dlPFC, and mechanisms purportedly underpinning BED psychopathology – behavioral approach and inhibition. Specifically, we leveraged the Adolescent Brain Cognitive Development (ABCD) Study (Casey et al., Reference Casey, Cannonier, Conley, Cohen, Barch and Heitzeg2018) to undertake the first known assessment of sex differences in GMD in pre-adolescent children with DSM-5 diagnosed BED. Since no studies have previously assessed sex differences in the neurobiology of BED, no a priori hypotheses were developed, and as such, the study was exploratory in nature.
Methods
Study sample
The ABCD Study is a large, diverse, and prospective cohort study of brain development and health throughout adolescence. The ABCD 3.0 release from baseline (Year 0) consists of 11 875 pre-adolescent children aged 9–10 years collected in 2016–2018, recruited from 21 sites around the U.S. Multi-stage probability sampling was employed to ensure, to the best extent possible, that the sample reflected an unbiased representation of the U.S. population and its major subpopulations (Garavan, Bartsch, Conway, & Zahs, Reference Garavan, Bartsch, Conway, Decastro, Goldstein, Heeringa and Zahs2018). With 21 primary recruitment sites, study recruitment leveraged a probability sampling of schools within the defined catchment area for each site, and eligible children in each sample school. Full details of the study sample, recruitment process, exclusion criteria, procedures, and measures have been previously reported (Barch et al., Reference Barch, Albaugh, Avenevoli, Chang, Clark, Glantz and Sher2018; Garavan et al., Reference Garavan, Bartsch, Conway, Decastro, Goldstein, Heeringa and Zahs2018). From this parent dataset, we extracted and analyzed data from 71 children diagnosed with BED and 74 non-psychiatric control children who were matched on age, BMI, and developmental maturation. Centralized institutional review board (IRB) approval was obtained from the University of California, San Diego. Study sites obtained approval from their local IRBs. Caregivers provided written informed consent and each child provided written assent.
Control group participants matching
Control group participants were matched according to BMI and pubertal development. The mean and standard deviation for these variables were calculated for the BED groups. Subsequently, participants without BED were extracted from the parent data set if they had a (i) BMI, (ii) self-reported PDS score and (iii) parent-reported PDS score within half of one standard deviation of the mean of the BED group.
Measures
Diagnostic screening
Parents/caregivers completed the eating disorder module of the Kiddie Schedule for Affective Disorders and Schizophrenia (KSADS-5) (Kaufman, Birmaher, & Brent, Reference Kaufman, Birmaher and Brent1997), assessing frequency, duration, and associated distress of their child's eating behavior. Parent/caregiver reports were self-administered via an online platform, which demonstrates excellent concordance with clinician administered KSADS interviews (Townsend et al., Reference Townsend, Kobak, Kearney, Milham, Andreotti, Escalera and Kaufman2020), and all diagnoses were made according to DSM-5 criteria for BED (American Psychiatric Association, 2013).
Pubertal Development Scale (PDS)
A self-reported and parent-reported measure of pubertal status (Herting et al., Reference Herting, Uban, Gonzalez, Baker, Kan, Thompson and Sowell2021; Petersen, Crockett, Richards, & Boxer, Reference Petersen, Crockett, Richards and Boxer1988), frequently used as a measure of developmental maturation when studying brain function and structure (Blakemore, Burnett, & Dahl, Reference Blakemore, Burnett and Dahl2010; Goddings, Beltz, Peper, Crone, & Braams, Reference Goddings, Beltz, Peper, Crone and Braams2019), was used in the present study to control for differential rates of maturation. Youth self-reported Pubertal Development Scale (PDS) scores were used in all analyses. While data generally suggests high internal consistency across self- and parent-reported versions of the PDS (Koopman-Verhoeff, Gredvig-Ardito, Barker, Saletin, & Carskadon, Reference Koopman-Verhoeff, Gredvig-Ardito, Barker, Saletin and Carskadon2020), additional evidence suggests that self-reported pubertal development correlates well with physician assessment, and hormonal assessment (Shirtcliff, Dahl, & Pollack, Reference Shirtcliff, Dahl and Pollack2009). For purposes of ensuring the most rigorous control participant matching, control participants were matched based on both self-reported and parent-reported PDS score. The correlation between child and parent reported PDS range from 0.2 to 0.5 ABCD Release 1.0 of the ABCD dataset (Barch et al., Reference Barch, Albaugh, Avenevoli, Chang, Clark, Glantz and Sher2018).
Behavioral Inhibition System/Behavioral Activation System Scale (BIS/BAS scale)
The BIS/BAS scale is a 24-item widely-used measure of self-reported tendencies towards goal-oriented hedonic pursuits, and behavioral inhibition, which demonstrates good psychometric properties (Carver & White, Reference Carver and White1994). Parent-reported BIS/BAS scores were used in analyses. Preliminary assessment of internal consistency of BIS/BAS subscale scores in Release 1.0 of the ABCD dataset yield acceptable alphas, ranging from 0.62 to 0.78.
Body mass index
BMI was calculated based on the average of two-to-three measured heights and weights (BMI = weight/height2) by research staff.
MRI data acquisition and preprocessing
Data and preprocessing was identical to the previous study using this cohort (Murray et al., Reference Murray, Duval, Balkchyan, Cabeen, Nagata, Toga and Jann2022b): Structural T1-weighted images were collected on Siemens Prisma (TR/TE = 2500/2.88 ms, 176 slices, 256 × 256 matrix size, voxel size 1 × 1 × 1 mm3, FA = 8°), Philips Achieva (TR/TE = 6.31/2.9 ms, 225 slices, 256 × 240 matrix size, voxel size 1 × 1 × 1mm3, FA = 8°) and GE MR750 (TR/TE = 2500/2 ms, 208 slices, 256 × 256 matrix size, voxel size 1 × 1 × 1mm3, FA = 8°) scanner platforms across 21 different sites within the United States of America. A Kolmogorov-Smirnov test revealed no statistical difference between scan-site distribution in the current BED and CONT groups (K = 0.094, p = 0.872). Individual T1 weighted images were segmented into different tissue types based on tissue probability primers for gray matter, white matter and CSF using SPM12 and the Computational Anatomy Toolbox (CAT12). Tissue maps were normalized to MNI space using the standard CAT12 DARTEL and Geodesic Shooting templates, which are derived from 555 healthy control subjects, smoothed with an 8 mm FWHM Gaussian Kernel and proportionally scaled to correct for total intracranial volume (TIV).
Statistical analyses
We employed a 2 × 2 ANCOVA with factors gender (male/female) and diagnosis (BED/CONT) including BMI and YPDS as covariates to analyze group differences. Of specific interest were the effect of gender and the interaction effect between gender and diagnostic group, as the effect of diagnostic group alone has been previously reported by our group (Murray et al., Reference Murray, Duval, Balkchyan, Cabeen, Nagata, Toga and Jann2022b). Post-hoc analyses to elucidate differences in average GMD between females and males within the BED group were performed by a 2-sample, 2-sided t test including covariates for pubertal status and BMI. Comparisons of average GMD were also performed between females and males within the control (CONT) group. Statistical significance was defined at p < 0.05 and correction for multiple comparison was performed by Threshold Free Cluster Enhancement (TFCE) with 5000 iterations (Smith & Nichols, Reference Smith and Nichols2009). Results are displayed on surface projections. Anatomical labels were obtained based on the Destrieux atlas implemented in CAT12 (Destrieux, Fischl, Dale, & Halgren, Reference Destrieux, Fischl, Dale and Halgren2010). Additionally, for specific ROIs which showed gender differences in BED we calculated effect sizes using eta2.
Brain-behavior associations
Post hoc partial correlation analysis was undertaken to assess the relationship between average regional voxel-based morphometry (VBM) metrics in a large cluster showing significant group differences in the dlPFC, and markers of behavioral inhibition and approach tendencies. We assessed this region of interest owing to (i) the noted role of the dlPFC in mechanisms putatively linked to the psychopathology of BED, (ii) our group's recent data illustrating altered GMD and functional connectivity in children with BED, which is linked to altered behavioral approach, and (iii) evidence of large sex differences in dlPFC GMD in children with BED. Additionally, we performed an analogous analysis within the control group in a smaller cluster within the dlPFC.
Results
Sample demographics
We identified 71 children (33 females; 38 males) with BED diagnoses based on DMS-5 criteria, and 74 non-psychiatric control children (38 females; 36 males), from the baseline visit of the ABCD study. Sex differences were not observed for age (p = 0.419), BMI (p = 0.962), youth reported pubertal development (p = 0.072), or total Behavioral Inhibition Scale score (p = 0.209). Girls' parent reported pubertal development score was greater (p = 0.004), whereas boys demonstrated greater Behavioral Activation Scale score (p = 0.037). Table 1 illustrates a detailed characterization of the sociodemographic characteristics of our study sample of children with BED, in addition to the characteristics of an age, BMI, and developmentally matched control group.
Table 1. An overview of the demographic characteristics of those with BED and the control group, delineated by sex

Voxel-based morphometry
Global volumes: between-group comparison
TIV significantly differed between females and males with BED [t(df = 69) = −3.03, p = 0.0035] and between females and males in the CONT group [t(df = 72) = −4.95, p < 0.0001]. Each individual's data was proportionally scaled to their corresponding TIV value to correct for this confounding factor.
Whole Brain Analysis: Between-Group Comparison: The ANCOVA main effect for gender revealed widespread significant differences in bilateral temporal, occipital, parietal and frontal lobes (See online Supplementary Fig. S1 and Table 2). The ANCOVA interaction effect for gender × diagnostic group did reveal three localized areas with significant interaction effects: in superior frontal gyrus, precuneus, and lingual sulcus (See online Supplementary Fig. S1 and Table 2). Post-hoc t tests for gender differences within BED and CONT groups individually were performed to elucidate the directions of effect (See Tables 3 & 4 and Fig. 1). VBM analysis revealed overlapping regions when comparing sex differences within the BED group and the CONT group (Fig. 1). Specifically, females in both the BED and CONT groups showed significantly greater GMD in the left postcentral gyrus, left precentral gyrus, and right superior frontal gyrus (SFG) (females > males). Preadolescent males in both the BED and CONT group, on average, showed greater GMD in the right middle temporal gyrus (MTG), right calcarine sulcus, and the cerebellum (males > females).
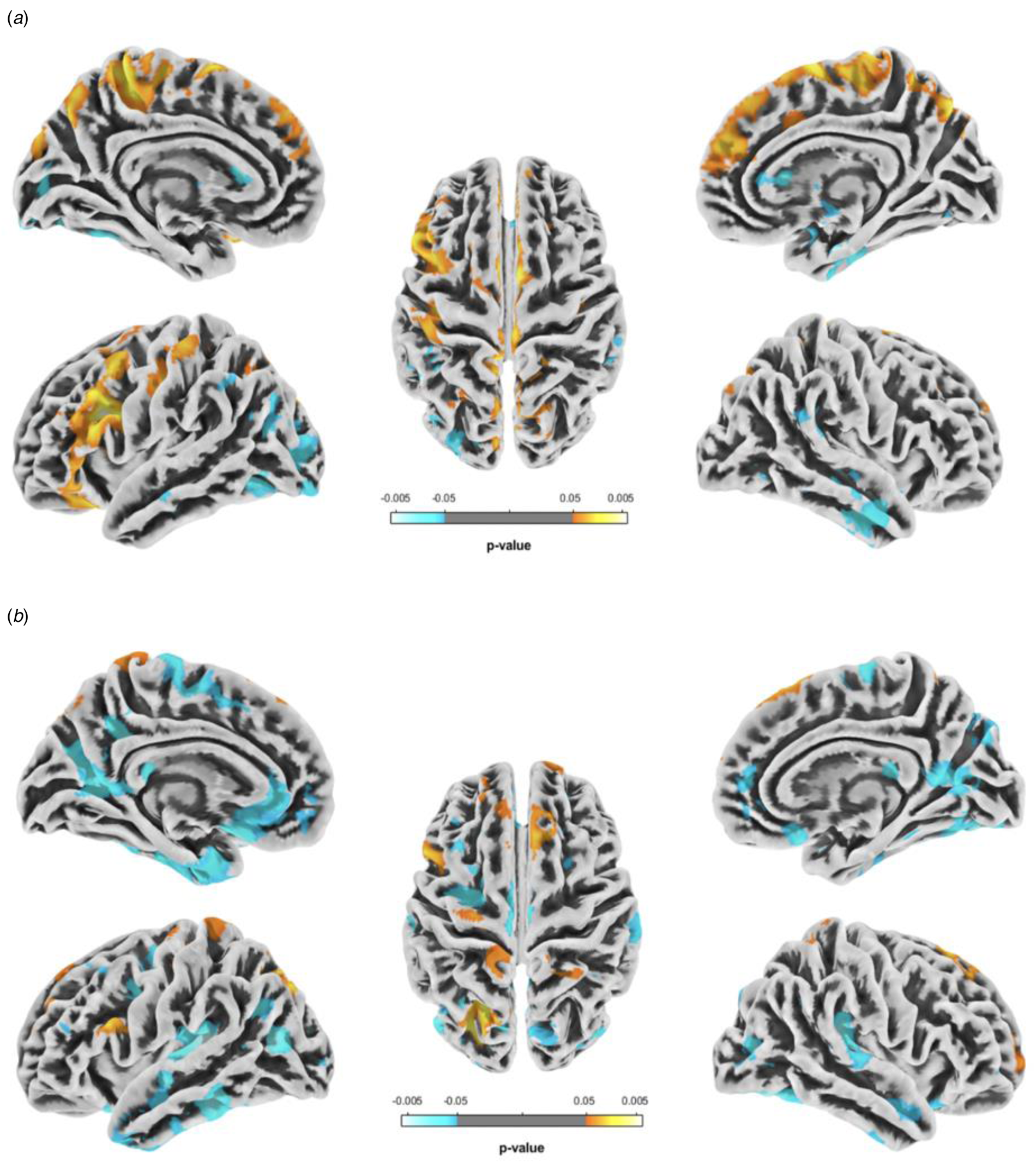
Fig. 1. VBM analyses illustrating regional sex differences in GMD among pre-adolescent girls with BED, relative to pre-adolescent boys with BED (a), and matched control girls without BED relative to matched control boys without BED (b). Hot colors indicate areas where pre-adolescent girls present with increased GMD as compared to pre-adolescent boys, while cool colors highlight areas where boys have increased GMD compared to girls.
Table 2. Anatomical location of cortical areas showing significant ANVOCA effects for main effects gender and for interaction effect gender × diagnostic group

Table 3. Anatomical location of cortical areas showing significant greater GMD among preadolescent males (N = 38) and females (N = 33) with BED

Table 4. Anatomical location of cortical areas showing significant greater GMD among typically developing preadolescent male (N = 36) and female (N = 38) controls

VBM analysis within the BED group revealed significantly greater GMD in preadolescent females in the left intraparietal sulcus (IPS), bilateral middle frontal gyrus (MFG), right central sulcus (CS), right anterior midcingulate cortex, and right precuneus (females > males) (Fig. 1a). In addition, preadolescent males with BED on average showed larger gray matter volumes in bilateral middle temporal gyrus (MTG), bilateral supramarginal gyrus (SMG), left inferior occipital gyrus (IOG), right middle occipital gyrus (MOG), right lingual gyrus, and right putamen (Pu) (males > females).
In contrast, preadolescent females without BED demonstrated greater GMD in the left IFG, left CS, left precuneus, bilateral superior parietal lobule (SPL), and right transverse frontopolar gyri (TFPG) (females > males). Preadolescent males without BED on average showed greater GMD in the left inferior temporal gyrus (ITG), bilateral superior temporal gyrus (STG), left middle frontal gyrus (MFG), bilateral superior frontal gyrus (SFG), left superior frontal sulcus (SFS), left MOG, right middle occipital sulcus (MOS), right inferior occipital gyrus and sulcus, right inferior parietal lobule (IPL), right orbital gyri, right cuneus, and right thalamus (Fig. 1b) (males > females).
In sum, girls with BED uniquely demonstrate diffuse clusters of significantly greater GMD in the (i) left dlPFC (eta2 = 0.082), (ii) bilateral dmPFC (eta2 = 0.061), (iii) bilateral primary motor and somatosensory cortex (eta2 = 0.092) and (iv) bilateral precuneus (eta2 = 0.068).
Brain-behavior associations
There were no significant associations between dlPFC GMD and either behavioral approach (r = −0.2; p = 0.245) or inhibition (r = −0.16; p = 0.2345) among boys with BED. Among girls with BED, no relationship emerged between dlPFC and behavioral inhibition (r = 0.3; p = 0.881), although a significant and negative correlation was observed for behavioral approach (r = −0.5; p = 0.004) (Fig. 2). Furthermore, for the control group we did not find any significant relation between dlPFC GMD and approach (female r = −0.08; p = 0.656/male r = −0.01; p = 0.941) or inhibition (female r = −0.02; p = 0.893/male r = −0.08; p = 0.643) (See online Supplementary Fig. S2).

Fig. 2. Region of interest analyses illustrating sex differences in GMD in the left dlPFC, among those with BED. (a) Box plot displaying mean and individual subject data points for both female and male participants. (b) Association or dlPFC-GMD with behavioral approach and inhibition scores assessed by BAS and BIS for pre-adolescent girls with BED, and (c) the associations of dlPFC-GMD with behavioral approach and inhibition scores in pre-adolescent boys with BED. Models in B and C have been adjusted for TIV and pubertal status. A significant association was found only in girls with BED where dlPFC-GMD negatively correlated with total BAS score.
Discussion
A nascent body of evidence has suggested nuanced sex differences in the psychopathology of BED, although potential sex differences in the neurobiology of BED has remained unknown. Here we present the first known assessment of sex differences in gray matter characteristics in BED, via VBM among a sample of 9–10-year-old children who met DSM-5 criteria for BED. In tandem with our assessment of gray matter characteristics of boys and girls with BED, we assessed sex differences in GMD among weight and developmentally-matched male and female controls, to ensure that any sex differences among those with BED were not representations of broader sexually dimorphic brain development among children. Results suggest a series of unique sex differences in GMD among those with BED, which were not evident among control children. Specifically, the most notable findings among those with BED suggested that girls with BED uniquely demonstrate diffuse clusters of greater GMD in the (i) left dlPFC, (ii) bilateral dmPFC, (iii) bilateral primary motor cortex, (iv) bilateral precuneus, and (v) bilateral primary somatosensory cortex. Moreover, assessment of sex differences in brain-behavioral associations revealed a discrepant relationship between left dlPFC GMD and markers of behavioral approach among girls and boys with BED, respectively.
Broadly, our findings illustrating several diffuse clusters of greater GMD in girls with BED, relative to boys with BED, are noteworthy in that they diverge from a robust body of evidence documenting sex differences in brain development during childhood and adolescence (Kaczkurkin, Raznahan, & Satterthwaite, Reference Kaczkurkin, Raznahan and Satterthwaite2019). Typically, brain maturation occurs developmentally earlier in girls relative to boys, and the earlier onset of arborization, synaptic pruning and myelination of long-range fiber bundles among girls is reflected in greater reductions in GMD during childhood and adolescence (Gennatas et al., Reference Gennatas, Avants, Wolf, Satterthwaite, Ruparel, Ciric and Gur2017). Indeed, this pattern was evident among our control group, where we observed a preponderance of regions of reduced GMD among girls, relative to matched control boys. These antonymic sex differences in girls with BED pose intriguing questions around the interaction between early onset BED and cortical maturation in girls.
While speculative, one interpretation of these findings may implicate gonadal sex hormones. Animal studies have demonstrated a reliably higher proclivity towards binge eating susceptibility among females in the context of palatable foods (Babbs, Wojnicki, & Corwin, Reference Babbs, Wojnicki and Corwin2012; Carlin et al., Reference Carlin, McKee, Hill-Smith, Grissom, George, Lucki and Reyes2016; Freund, Thompson, Norman, Einhorn, & Andersen, Reference Freund, Thompson, Norman, Einhorn and Andersen2015; Hardaway et al., Reference Hardaway, Jensen, Kim, Mazzone, Sugam, Diberto and Kash2016). Importantly, sex differences in animal models of binge eating become more pronounced alongside increasingly severe and frequent binge eating, which mirrors sex differences in more severe presentations of binge eating in human populations (Klump, Culbert, & Sisk, Reference Klump, Culbert and Sisk2017). These effects have been putatively attributed to the impact of gonadal hormones, both in pre- and peri-natal periods, and subsequently during puberty (Klump et al., Reference Klump, Culbert and Sisk2017). Specifically, evidence from animal and human populations has illustrated that elevations in estrogen and progesterone portend elevations in binge eating tendencies (Klump et al., Reference Klump, Culbert and Sisk2017). In concert, estrogen has profound neuroprotective properties in a multitude of settings, dually bolstering the prevention of gray matter loss and facilitating increases in gray matter volume (Albert et al., Reference Albert, Hiscox, Boyd, Dumas, Taylor and Newhouse2017; Lord, Engert, Lupien, & Pruessner, Reference Lord, Engert, Lupien and Pruessner2010; MacKenzie-Graham et al., Reference MacKenzie-Graham, Rinek, Avedisian, Morales, Umeda, Boulat and Voskuhl2012). However, gray matter loss is an essential component of typical neurodevelopmental maturation, where synaptic pruning and gray matter arborization are characteristic. To this end, estradiol in the developing brain has a negative correlation with GMD – in that higher estradiol is associated with lower GMD (Brouwer et al., Reference Brouwer, Koenis, Schnack, van Baal, van Soelen, Boomsma and Hulshoff Pol2015). While both estrogen and progesterone – which has an antagonistic effect on estrogen – are elevated in those with BED, these findings pose the intriguing question around whether the sex hormones linked to greater binge eating are also linked to altered synaptic pruning and gray matter arborization. While altered synaptic pruning and arborization would impact the functional connectivity between gray matter structures, recent evidence has illustrated pervasive functional dysconnectivity in pre-adolescent children with BED (Murray et al., Reference Murray, Alba, Duval, Nagata, Cabeen, Lee and Jann2022a), which persists into adulthood (Haynos et al., Reference Haynos, Camchong, Pearson, Lavender, Mueller, Peterson and Lim2021). However, the delineation of potential sex differences in functional connectivity would be critical in advancing this line of inquiry.
Alongside the broad view of elevated GMD in girls with BED, specific clusters are noteworthy. A large cluster of elevated GMD was evident in the left dlPFC of girls with BED. The dlPFC has been reliably implicated in inhibitory control, and is thought be a central node of this network (Anderson & Weaver, Reference Anderson, Weaver and Squire2009; Constantinidis & Luna, Reference Constantinidis and Luna2019; Penolazzi, Stramaccia, Braga, Mondini, & Galfano, Reference Penolazzi, Stramaccia, Braga, Mondini and Galfano2014). To that end, functional (Celone, Thompson-Brenner, Ross, Pratt, & Stern, Reference Celone, Thompson-Brenner, Ross, Pratt and Stern2011; Murray et al., Reference Murray, Alba, Duval, Nagata, Cabeen, Lee and Jann2022a) and structural (Murray et al., Reference Murray, Duval, Balkchyan, Cabeen, Nagata, Toga and Jann2022b) perturbations in the dlPFC have been documented in those with BED, although no studies to date have assessed sex differences. Our findings suggest that the altered dlPFC GMD observed in children with BED (Murray et al., Reference Murray, Duval, Balkchyan, Cabeen, Nagata, Toga and Jann2022b) may be skewed towards girls. Notably, and in keeping with the noted role of the dlPFC in emotional regulation (Etkin, Büchel, & Gross, Reference Etkin, Büchel and Gross2015; Golkar et al., Reference Golkar, Lonsdorf, Olsson, Lindstrom, Berrebi, Fransson and Öhman2012), our findings may offer preliminary insights on the greater proclivity of girls with BED to engage in binge eating in response to negative affect (Stice, Akutagawa, Gaggar, & Agras, Reference Stice, Akutagawa, Gaggar and Agras2000), and report stronger negative affect following binge episodes (DiGioacchino, Sargent, Sharpe, & Miller, Reference DiGioacchino, Sargent, Sharpe and Miller1999).
Interestingly, our behavioral findings suggest a significantly lower tendency towards behavioral approach among girls with BED. Assessment of brain-behavioral relationships suggests that this lower behavioral approach among girls with BED may be uniquely linked to the perturbations in dlPFC morphometry also observed in girls with BED. Specifically, we observed a significant and uniquely negative correlation between dlPFC GMD and behavioral approach among girls with BED, insofar as lower behavioral approach scores were significantly associated with greater elevations in dlPFC GMD. This relationship was not observed among boys with BED or girls without BED in the present study (see online Supplementary Fig. S2), or among carefully matched control children of either sex in a previous study (Murray et al., Reference Murray, Duval, Balkchyan, Cabeen, Nagata, Toga and Jann2022b). This finding may have important implications for emerging neuromodulatory treatments, where for instance, the left dlPFC has been identified as a target for transcranial magnetic stimulation (TMS) for BED. Existing TMS trials for BED, alongside focusing on the left dlPFC as a target, have also focused largely on female samples (Gay et al., Reference Gay, Jaussent, Sigaud, Billard, Attal, Seneque and Guillaume2016; Maranhão et al., Reference Maranhão, Estella, Cury, Amigo, Picasso, Berberian and Claudino2015), with a unified focus on reducing behavioral approach by palatable food cues. Our findings suggest that the investigation of sex differences in the efficacy of left dlPFC-centered TMS may be a critical next step as TMS treatments for BED advance.
Diffuse and bilateral elevations in GMD were also observed in dorsomedial prefrontal cortices (dmPFC) among girls with BED. Broadly, evidence has illustrated altered medial PFC activity in those with BED when assessing inhibitory control (Balodis et al., Reference Balodis, Molina, Kober, Worhunsky, White, Sinha and Potenza2013) and reward processing (Balodis et al., Reference Balodis, Grilo, Kober, Worhunsky, White, Stevens and Potenza2014), which is linked to the persistence of binge episodes following treatment (Balodis et al., Reference Balodis, Grilo, Kober, Worhunsky, White, Stevens and Potenza2014). With specific regards to the dmPFC, TMS to this region has been linked to the rapid remission of binge-type symptoms in bulimia nervosa (Downar, Sankar, Giacobbe, Woodside, & Colton, Reference Downar, Sankar, Giacobbe, Woodside and Colton2012), although treatment effects are diluted among those with impoverished pre-treatment connectivity between the dmPFC, and OFC, insula and temporal pole, respectively (Dunlop et al., Reference Dunlop, Woodside, Lam, Olmsted, Colton, Giacobbe and Downar2015). In concert, animal studies illustrate altered neuronal signaling in the dmPFC of rats with binge eating, suggesting that ‘binge resistance’ may be dependent upon mPFC circuitry (Corwin et al., Reference Corwin, Wojnicki, Zimmer, Babbs, McGrath, Olivos and Hayes2016). Cumulatively, these findings may offer important insights into the greater association between impulsivity and lifetime BED among females, relative to males (Lee-Winn, Townsend, Reinblatt, & Mendelson, Reference Lee-Winn, Townsend, Reinblatt and Mendelson2016).
We also noted bilaterally elevated in GMD in the precuneus of girls with BED. The precuneus is typically associated with processes around self- and visuospatial-awareness (Cavanna & Trimble, Reference Cavanna and Trimble2006; Schott et al., Reference Schott, Wüstenberg, Lücke, Pohl, Richter, Seidenbecher and Richardson-Klavehn2019), although aberrant precuneus activity has consistently been noted during tasks of body evaluation among those with an array of eating disorders (Gaudio & Quattrocchi, Reference Gaudio and Quattrocchi2012; Lee et al., Reference Lee, Ran Kim, Ku, Lee, Namkoong and Jung2014). While the neural correlates of body image among those with BED remains understudied, studies assessing sex differences in body self-evaluation found that following exposure to overweight images, females demonstrated greater precuneus activity than males when evaluating their body (Owens, Allen, & Spangler, Reference Owens, Allen and Spangler2010). Among those with BED, women typically report greater levels of body dissatisfaction (Grilo & Masheb, Reference Grilo and Masheb2005a, Reference Grilo and Masheb2005b; Grilo, Masheb, Brody, Burke-Martindale, & Rothschild, Reference Grilo, Masheb, Brody, Burke-Martindale and Rothschild2005), and among community populations, preoccupation with weight and shape is a reliably potent predictor of binge eating behavior in girls (Mitchison et al., Reference Mitchison, Hay, Griffiths, Murray, Bentley, Gratwick-Sarll and Mond2017). Studies assessing sex differences in body self-evaluation found that following exposure to overweight images, females demonstrated greater precuneus activity than males when evaluating their body (Owens et al., Reference Owens, Allen and Spangler2010). Our findings noting bilateral elevations in precuneal GMD among girls with BED may offer insights into a possible biological basis for the greater relationship between body image concerns and disordered eating in girls with BED.
Lastly, we also noted bilateral elevations in GMD in primary motor and sensory cortices among girls with BED. While a recent gender-conflated assessment of regional GMD abnormalities among children with BED suggested small clusters of elevated GMD in pre- and postcentral gyri relative to tightly matched control children (Murray et al., Reference Murray, Duval, Balkchyan, Cabeen, Nagata, Toga and Jann2022b), the delineation of sex differences suggests that these differences are likely driven by diffusely elevated GMD in these regions among girls with BED. Previous studies have noted bilateral elevations in neural activity in precentral gyri among girls with binge-type eating disorders (Lock, Garrett, Beenhakker, & Reiss, Reference Lock, Garrett, Beenhakker and Reiss2011) during tasks of inhibitory control. Additional studies of adults with binge-type eating disorders have illustrated altered activity in precentral gyri in response to food cues (Brooks et al., Reference Brooks, O'Daly, Uher, Friederich, Giampietro, Brammer and Campbell2011). Similarly, studies of BN in women suggest diminished activity in postcentral gyri during tasks of food-related behavioral inhibition (Skunde et al., Reference Skunde, Walther, Simon, Wu, Bendszus, Herzog and Friederich2016). However, these studies have predominantly included women with bulimia nervosa, which, while inclusive of binge episodes, is characterized by discrepant core symptoms which cannot be seamlessly extrapolated to BED. Our findings underscore the importance of delineating findings by sex.
Strengths of the present study include the relatively large sample size, which permitted the first known assessment of sex differences of the neurobiology of BED. In addition, our focus on pre-adolescent BED offers important insights into gray matter development among the broader backdrop of known sexually dimorphic patterns in brain development. Moreover, the assessment of sex differences in GMD among tightly matched controls allowed the isolation of sex differences specific to BED. However, several limitations of the study are worthy of discussion. Firstly, clinical observations suggest that sex differences in BED may be less pronounced during pre-adolescence, but diverge more markedly following puberty. Specifically, while binge eating remains relatively stable among boys throughout puberty and into adulthood, girls typically report greater levels of binge eating (Culbert et al., Reference Culbert, Sinclair, Hildebrandt, Burt, Sisk and Klump2016). This is reflected in the prevalence of BED in boys and girls in our sample of 9–10-year-old children. An important question therefore relates to the assessment of neurobiological sex differences as clinical sequalae start to diverge, and as the developmental trajectory of gray matter morphometry evolves throughout adolescence into adulthood. Additionally, and owing to the characteristics of the parent ABCD dataset, no dimensional measure of BED symptomatology was included, which precluded assessment of the extent to which the observed sex differences in GMD differentially relate to symptom severity. Moreover, and while based on DSM-5 criteria, diagnoses in this instance were drawn from parental observations. With noted discrepancies between child and parent reports of BED symptomatology, this is a limitation. In addition, the sample size was relatively modest, and appropriate caution is advised when interpreting findings. Importantly, the presence and impact of psychiatric comorbidities was not assessed in the present study, and should be expanded upon in future studies. Lastly, and in keeping with the limitations of the parent ABCD dataset, hydration and hunger status were not assessed prior to scanning in the present study.
Notwithstanding, our findings present novel data around sex differences in gray matter morphometry in the developing brain of children with BED, providing evidence of antonymic patterns of brain development between girls with and without BED, relative to boys. Future research may extend these findings by assessing possible sex differences in other domains of brain structure and function among children with BED. Importantly, these data may raise the possibility of sex-specific effects of brain-based treatments which target structures with disparate characteristics between boys and girls with BED, respectively.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0033291722003269
Acknowledgments
None.
Financial support
SBM is supported by the Della Martin Endowed Professorship, and the National Institute of Mental Health (K23MH115184). J.M.N. was funded by the National Heart, Lung, and Blood Institute (K08HL159350) and the American Heart Association (CDA34760281). RPC is supported by the Chan Zuckerberg Initiative (2020-225670). SJS is supported by Astellas.
SBM receives royalties from Oxford University Press, Routledge, and Springer. JMN receives royalties from Springer. SJS is a Consultant to Zynerba, and an Advisor to Skyland Trail.
Additional information
The ABCD Study was supported by the National Institutes of Health and additional federal partners under award numbers U01DA041022, U01DA041025, U01DA041028, U01DA041048, U01DA041089, U01DA041093, U01DA041106, U01DA041117, U01DA041120, U01DA041134, U01DA041148, U01DA041156, U01DA041174, U24DA041123, and U24DA041147. A full list of supporters is available at https://abcdstudy.org/nihcollaborators. A listing of participating sites and a complete listing of the study investigators can be found at https://abcdstudy.org/principal-investigators.html. ABCD consortium investigators designed and implemented the study and/or provided data but did not necessarily participate in the analysis or writing of this report.
Conflict of interest
The authors all declare that they have no competing interests.








