



1. Introduction
The Turonian-age Bissekty Formation in the central Kyzylkum Desert of Uzbekistan has yielded one of the most diverse assemblages of early Late Cretaceous terrestrial and freshwater vertebrates found to date. The dinosaurs from this assemblage include representatives of various clades of non-avian theropods: a tyrannosauroid (Timurlengia euotica, Brusatte et al. Reference Brusatte, Averianov, Sues, Muir and Butler2016); dromaeosaurids (Itemirus medullaris and a large-bodied form, Sues & Averianov, Reference Sues and Averianov2014); troodontids (Urbacodon sp., Averianov & Sues, Reference Averianov and Sues2007); alvarezsaurids (Dzharaonyx etsi, Averianov & Sues, Reference Averianov and Sues2022); an unnamed ornithomimid; at least two unnamed taxa of therizinosauroids; and a caenagnathid (Caenagnathasia martinsoni, Sues & Averianov, Reference Sues and Averianov2015). In addition, there are two form taxa for distinctive small theropod teeth, Richardoestesia americana and Paronychodon asiaticus (Sues & Averianov, Reference Sues and Averianov2013; Averianov & Sues, Reference Averianov and Sues2019). Recently, Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) described what they considered a neovenatorid carcharodontosaurian, Ulughbegsaurus uzbekistanensis, which will be further discussed in this paper.
Although almost invariably dissociated, the bones and teeth of these dinosaurs are often exquisitely preserved and provide a wealth of anatomical information. At Dzharakuduk in the Navoi District of Uzbekistan, strata of the Bissekty Formation are widely exposed along an escarpment that extends from approximately 42° 06′ 22.60″ N, 62° 37′ 09.00″ E to 42° 05′ 44.22″ N, 62° 4′ 06.49″ E. The Bissekty Formation encompasses an up to 80 m thick succession of poorly lithified, medium-grained and cross-bedded fluvial sandstones and several laterally extensive, clast-supported intraformational conglomerates (Redman & Leighton, Reference Redman and Leighton2009). The Bissekty Formation is underlain by the Dzheirantui Formation and overlain by the Aitym Formation, both of which were deposited in marginal- or shallow-marine environments. Based on the fieldwork by the late Christopher King (pers. comm.), the Dzheirantui Formation can be dated as latest early Turonian based on the co-occurrence of two taxa of inoceramid bivalves, Mytiloides aff. M. labiatus and Mytiloides subhercynicus. The Meshekeli Member of the Aitym Formation is late Turonian primarily based on the presence of the inoceramid Mytiloides incertus. Thus, the age of the Bissekty Formation is middle Turonian (Averianov, Reference Averianov2010).
Most bones and teeth of non-avian dinosaurs from the Bissekty Formation have been recovered by surface collecting. As expected in a fluvial depositional setting, many skeletal remains show signs of postmortem transport. For some bones, preservation was further adversely affected by prolonged surface exposure in the harsh present-day desert climate.
Institutional abbreviations: BYU – Museum of Paleontology, Department of Geological Sciences, Brigham Young University, Provo, Utah, USA; CCMGE – Chernyshev’s Central Museum of Geological Exploration, Saint Petersburg, Russia; UALVP – University of Alberta Vertebrate Palaeontology Lab, Edmonton, Alberta, Canada; UzSGM – State Geological Museum of the State Committee of the Republic of Uzbekistan on Geology and Mineral Resources, Tashkent, Uzbekistan; YPM – Peabody Museum of Natural History, Yale University, New Haven, Connecticut, USA; ZIN PH – Zoological Institute, Paleoherpetological Collection, Russian Academy of Sciences, Saint Petersburg, Russia.
2. Systematic palaeontology
DINOSAURIA Owen, Reference Owen1842
THEROPODA Marsh, Reference Marsh1881
PARAVES Sereno, Reference Sereno1997
DROMAEOSAURIDAE Matthew & Brown, Reference Matthew and Brown1922
EUDROMAEOSAURIA Longrich & Currie, Reference Longrich and Currie2009
Gen. et sp. indet.
Sues & Averianov (Reference Sues and Averianov2014) tentatively assigned all dromaeosaurid bones and teeth to Itemirus medullaris, which was originally named on the basis of an excellently preserved partial braincase (Kurzanov, Reference Kurzanov1976). Further review suggests most of the material represents a small- to medium-sized dromaeosaurid (based on fully closed sutures between the bones of the holotypic braincase and the closed neurocentral sutures on the available vertebrae), to which we apply the binomen Itemirus medullaris, and a very large dromaeosaurid that is the subject of the present study.
A complete left pedal phalanx II-2 ZIN PH 11/16 (Figs 1 and 2c) has a greatest length of 84.5 mm (correcting the measurement in Sues & Averianov, Reference Sues and Averianov2014). It was briefly described by Sues & Averianov (Reference Sues and Averianov2014). The proximal width of this phalanx is 41.3 mm and its distal width is 39.8 mm. The phalanx is proportionately shorter anteroposteriorly and wider transversely than the homologous bones in most known dromaeosaurids except in Achillobator giganticus from the Upper Cretaceous (Cenomanian–Santonian) Bayn Shire Formation of Mongolia (Perle et al. Reference Perle, Norell and Clark1999, pl. 13; length: 56.4 mm – PJ Currie, pers. comm.). By comparison, pedal phalanges II-2 of the up to 3 m long Deinonychus antirrhopus, from the Lower Cretaceous (Aptian–Albian) Cloverly Formation of Montana and Wyoming (Ostrom, Reference Ostrom1969), have lengths of up to 49.9 mm (Brusatte et al. Reference Brusatte, Vremir, Csiki-Sava, Turner, Watanabe, Erickson and Norell2013), and two pedal phalanges II-2 of Austroraptor cabazai from the Upper Cretaceous (Maastrichtian) Formation of Argentina are 58.1 and 58.8 mm long, respectively (Currie & Paulina Carabajal, Reference Currie and Paulina Carabajal2012). The phalanx ZIN PH 11/16 has a long, in dorsal/ventral view lobate proximoventral flange or ‘heel’ (Fig. 1c, d). The ventral surface of this heel is gently convex transversely rather than flat as in Deinonychus antirrhopus (YPM VP.005205). The presence of a well-developed proximoventral heel on pedal phalanx II-2 has been hypothesized as diagnostic for Eudromaeosauria (Longrich & Currie, Reference Longrich and Currie2009; Turner et al. Reference Turner, Makovicky and Norell2012). The proximal articular facet extends onto the heel and is asymmetrically divided by a median ridge into a lateral and a wider medial articular surface. Its dorsal margin forms a distinct median ‘lip’. As in Achillobator giganticus (Perle et al. 1999; Fig. 2b), the body of the phalanx is only slightly constricted in side view. By contrast, the body has a clearly defined ‘neck’ between the two articular ends in most other dromaeosaurids such as Deinonychus antirrhopus (YPM VP.005205; Fig. 2a). The distal end of ZIN PH 11/16 forms a grooved ginglymoid articular facet, which is semicircular (∼180°) in side view and narrower transversely than the proximal facet. As in other dromaeosaurids, its articular facet extends farther proximally onto the ventral surface than onto the dorsal surface of the bone. The lateral and medial surfaces of the distal end of the phalanx bear collateral ligament pits positioned posterodorsal to the geometrical centre of the ginglymus arc. The lateral pit is obscured by tightly adhering matrix whereas the medial pit is well-developed and visible even in dorsal view.

Fig. 1. Eudromaeosauria gen. et sp. indet., Bissekty Formation, left pedal phalanx II-2 (ZIN PH 11/16), in (a) lateral, (b) medial, (c) dorsal, (d) ventral, (e) proximal and (f) distal views. Scale bar = 3 cm. Abbreviations: clp, collateral ligament pit; pvh, posteroventral ‘heel’.
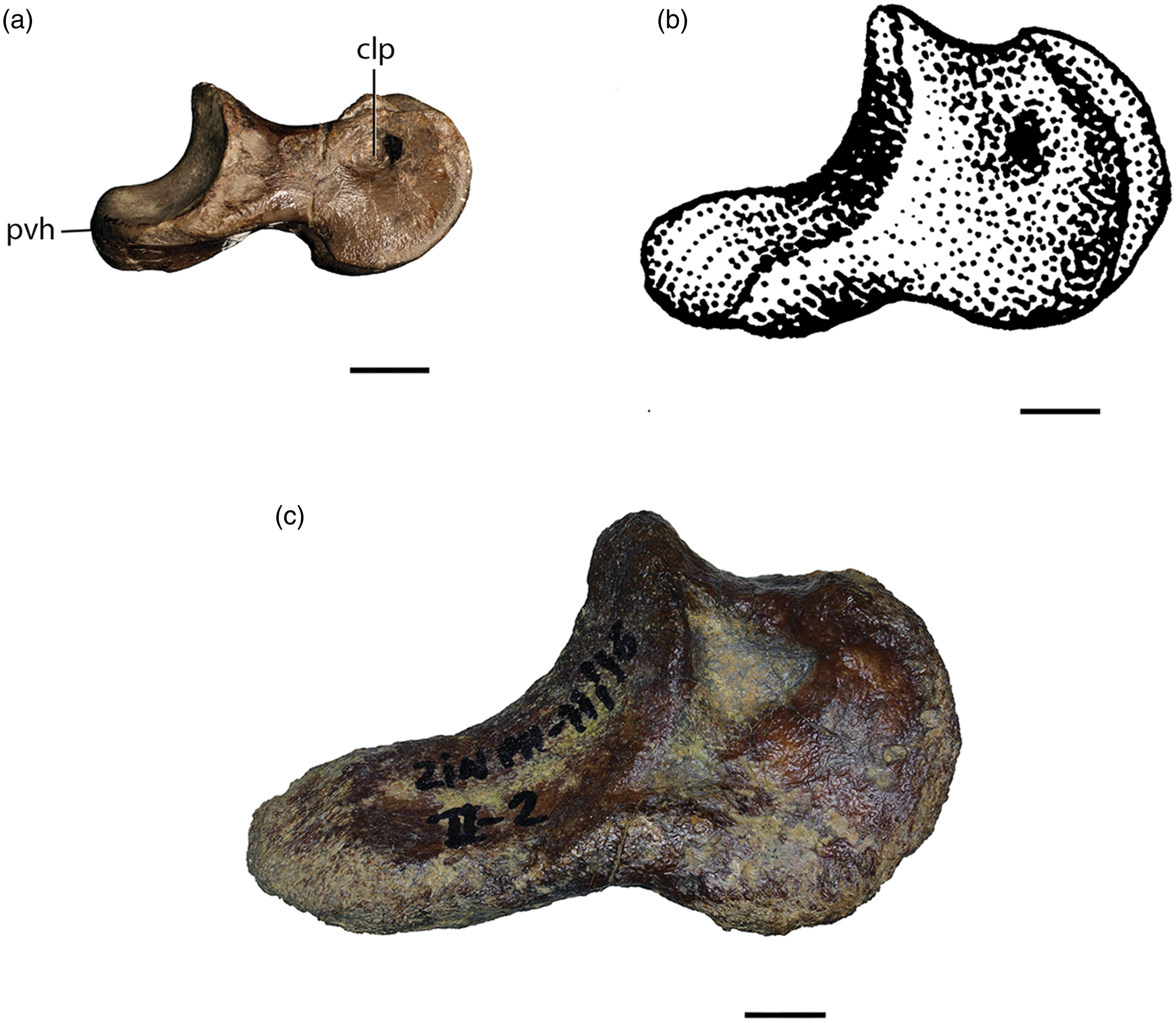
Fig. 2. Pedal phalanges II-2 of (a) Deinonychus antirrhopus (YPM VP.005205, reversed), (b) Achillobator giganticus and (c) ZIN PH 11/16, shown at the same scale for comparison. (a) Courtesy of DL Brinkman and (b) scanned and reversed from Perle et al. (1999, pl. 13). Scale bars each equal 1 cm. Abbreviations: clp, collateral ligament pit; pvh, posteroventral ‘heel’.
Sues & Averianov (Reference Sues and Averianov2014) described two fragments of the posterior ends of large maxillae, which closely resemble the postalveolar portions of the maxillae of Dromaeosaurus albertensis (Currie, Reference Currie1995). The more complete and better-preserved fragment of the posterior portion of a left maxilla (CCMGE 600/12457) is c. 150 mm long and preserves the posterior four alveoli and parts of two preceding the former. The posterior ramus of the maxilla extends posteriorly well behind the tooth row, which closely resembles the condition in dromaeosaurids such as Achillobator giganticus (Turner et al. Reference Turner, Makovicky and Norell2012) and Dromaeosaurus albertensis (Currie, Reference Currie1995) but is also present in some carcharodontosaurians (e.g. Acrocanthosaurus atokensis, Currie & Carpenter, Reference Currie and Carpenter2000).
3. Affinities of Ulughbegsaurus uzbekistanensis
Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) reported a fragment of a left maxilla of a large theropod from the Bissekty Formation, which they designated as the holotype of a new taxon of neovenatorid carcharodontosaurian, Ulughbegsaurus uzbekistanensis. Based on this specimen, the authors argued that the non-avian theropod assemblage from the Bissekty Formation resembles that from the Cenomanian Mussentuchit Member of the Cedar Mountain Formation in Utah (USA), in which the large neovenatorid Siats meekerorum (Zanno & Makovicky, Reference Zanno and Makovicky2013) occurred together with the diminutive tyrannosauroid Moros intrepidus (Zanno et al. Reference Zanno, Tucker, Canoville, Avrahami, Gates and Makovicky2019). If the identification of Ulughbegsaurus uzbekistanensis as a neovenatorid were substantiated, the Bissekty material would represent the geologically youngest example of a non-avian theropod assemblage in which a large carcharodontosaurian co-occurred with a medium-sized tyrannosauroid.
The holotype of Ulughbegsaurus uzbekistanensis (UzSGM 11-01-02) is a fragment of a left maxilla without erupted teeth (Fig. 3a, b) from Dzharakuduk. Here we reassess the phylogenetic relationships of this taxon based on the description and figures published by Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021).

Fig. 3. Partial maxillae of Ulughbegsaurus uzbekistanensis (holotype, UzSGM 11-01-02; a, b) and Utahraptor ostrommaysi (BYU 19965, reversed; c, d), each in (a, c) lateral and (b, d) medial views. (a, b) From Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) – CC BY 4.0. Scale bars each equal 5 cm. Abbreviations: amp, anteromedial process of maxilla; af, accessory fossa; aofe, margin of antorbital fenestra; aofo, antorbital fossa; idp, interdental plate; mfe, maxillary fenestra; nf, nutrient foramen; ‘pmfo’, ‘promaxillary fossa’; ps, palatal shelf; slf, supralabial foramen.
Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) listed several purported synapomorphies in support of the referral of U. uzbekistanensis to neovenatorid carcharodontosaurians. Firstly, they cited the rugose lateral surface of the holotypic maxilla. Their excellent photographs (Tanaka et al. Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021, figs 1a, 2a; Fig. 3a) show that the lateral surface of UzSGM 11-01-02 is badly eroded. The lateral surface of the maxilla fragment presents a distinctly fibrous internal structure, which is commonly observed on vertebrate bones after prolonged abrasion by sediment-laden flowing water (Behrensmeyer, Reference Behrensmeyer1978). Some of the bony fibres cross the margins of eroded neurovascular canals, creating the appearance of ‘ridging’. This pattern does not resemble the distinct ridging present on the lateral surface of the maxilla in undisputed carcharodontosaurians such as Carcharodontosaurus spp. (Brusatte & Sereno, Reference Brusatte and Sereno2007; Delcourt & Grillo, Reference Delcourt and Grillo2018; Ibrahim et al. Reference Ibrahim, Sereno, Varricchio, Martill, Dutheil, Unwin, Baidder, Larsson, Zouhri and Kaoukaya2020). Furthermore, we note that maxillae of various other theropods have rugose lateral surfaces with ridges and grooves extending from the neurovascular foramina, including Abelisauridae (Lamanna et al. Reference Lamanna, Martínez and Smith2002; Sampson & Witmer, Reference Sampson and Witmer2007), Dromaeosauridae (e.g. Dromaeosaurus albertensis, Utahraptor ostrommaysi, Fig. 3c) and Tyrannosauridae (e.g. Tarbosaurus bataar, Hurum & Sabath, Reference Hurum and Sabath2003). Thus, rugose lateral surfaces of the maxillae are not unique to carcharodontosaurians and, in the case of UzSGM 11-01-02, this feature is a taphonomic artefact.
Secondly, Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) considered the position of what they identified as a ‘promaxillary fossa’ on the anterior rim of the antorbital fossa as comparable to the condition in allosauroids. Their illustration (reproduced here as Fig. 3a) shows a small, round pit with irregular edges on the rounded anteroventral rim of the antorbital fossa, but it is unclear whether this is a genuine morphological feature or merely taphonomic damage similar to another, larger pit on the margin of the antorbital fossa. A small maxillary foramen is present in this position in Carcharodontosaurus spp. (Brusatte & Sereno, Reference Brusatte and Sereno2007) and a large maxillary fenestra in Neovenator salerii (Brusatte et al. Reference Brusatte, Benson and Hutt2008). However, the carcharodontosaurid Shaochilong moartuensis lacks such a foramen (Brusatte et al. Reference Brusatte, Chure, Benson and Xu2010). Given the variability in this feature even among carcharodontosaurians, its diagnostic value is questionable. Dromaeosaurids have promaxillary fenestrae of various sizes (Powers et al. Reference Powers, Fabbri, Doschak, Bhullar, Evans, Norell and Currie2022).
Thirdly, Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) cited the presence of fused, slightly rugose interdental plates in Ulughbegsaurus uzbekistanensis. The absence of distinct interdental plates is not restricted to most but not all allosauroids (Currie, Reference Currie1995) but is shared by abelisaurids (Lamanna et al. Reference Lamanna, Martínez and Smith2002; Sampson & Witmer, Reference Sampson and Witmer2007), the megalosauroid Torvosaurus spp. (Britt, Reference Britt1991; Hendrickx & Mateus, Reference Hendrickx and Mateus2014), dromaeosaurids (Currie, Reference Currie1995; Turner et al. Reference Turner, Makovicky and Norell2012) and troodontids (Currie, Reference Currie1987). By contrast, the tyrannosauroid Timurlengia euotica from the Bissekty Formation has distinct interdental plates (Averianov & Sues, Reference Averianov and Sues2012). Rugose interdental plates also occur in other non-avian theropods (e.g. Tarbosaurus bataar, Hurum & Sabath, Reference Hurum and Sabath2003).
Fourthly, Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) cited the presence of alveoli that are labiolingually narrower than mesiodistally long in Ulughbegsaurus uzbekistanensis as a similarity to carcharodontosaurians. However, dromaeosaurids (e.g. Saurornitholestes langstoni, UALVP 12339) also have alveoli that are distinctly more narrow labiolingually than long mesiodistally with ratios of labiolingual width to mesiodistal length of c. 0.5, comparable to the ratios cited by Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) for Ulughbegsaurus uzbekistanensis.
We could not identify a single feature that unambiguously supports referral of Ulughbegsaurus uzbekistanensis to neovenatorid carcharodontosaurians. Three of the purported synapomorphies cited by Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) are also shared by dromaeosaurids. In addition, some features like the sub-vertical contact between the premaxilla and maxilla and the smooth junction between the ventral margin of the antorbital fossa and the lateral surface of the maxilla ventral to this margin (Powers et al. Reference Powers, Fabbri, Doschak, Bhullar, Evans, Norell and Currie2022) are present in both Ulughbegsaurus uzbekistanensis and some dromaeosaurids such as Utahraptor ostrommaysi (Fig. 3c,d).
Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) presented a phylogenetic analysis that included only allosauroids and tyrannosauroids and excluded all other non-avian theropod clades. This selective taxon sampling assumed relationships a priori and did not allow a comprehensive assessment. Dromaeosaurids such as Achillobator giganticus from the Late Cretaceous (Cenomanian–Santonian) Bayn Shire Formation of Mongolia (Perle et al. 1999; Turner et al. Reference Turner, Makovicky and Norell2012) and Utahraptor ostrommaysi from the Lower Cretaceous (Barremian–Aptian) Upper Yellow Cat Member of the Cedar Mountain Formation of Utah (Kirkland et al. Reference Kirkland, Burge and Gaston1993) attained linear dimensions comparable to those of other large theropods. Indeed, Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) even made reference to ZIN PH 11/16 discussed here.
Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) referred the two fragments of maxillae described by Sues & Averianov (Reference Sues and Averianov2014) to Ulughbegsaurus uzbekistanensis, even though there is no anatomical similarity or overlap between the three specimens. They then reconstructed a remarkably long maxilla by combining the holotype and CCMGE 600/12457 (Tanaka et al. Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021, fig. S7) and based a high estimate of body size on this reconstruction. However, this reconstruction cannot be justified since there are no anatomical landmarks to associate the two fragments and determine their relative positions.
Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) referred CCMGE 600/12457 to Ulughbegsaurus uzbekistanensis based on the ‘beading’ along the ventral rim of the antorbital fossa. This ‘beading’ on the two jaw fragments is likely not an anatomical feature. For example, the ventral rim of the antorbital fossa on the lateral surface of the maxilla of the Utahraptor ostrommaysi is closely associated with a number of neurovascular foramina (Fig. 3c). Even slight surficial erosion would generate ‘beads’ from the slightly thickened bone around canals associated with these openings. This is clearly evident on the eroded lateral surface of the holotypic maxilla of Ulughbegsaurus uzbekistanensis (Tanaka et al. Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021, fig. 2b).
The structure of CCMGE 600/12457 resembles that in some carcharodontosaurians (e.g. Acrocanthosaurus atokensis, Currie & Carpenter, Reference Currie and Carpenter2000) but also that in dromaeosaurids such as Dromaeosaurus albertensis (Currie, Reference Currie1995). Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) cited the ‘approximately 20° ventral orientation at the jugal’ as a carcharodontosaurian synapomorphy but their assessment is based only on the referred fragment CCMGE 600/12457. Furthermore, this character-state is also shared by dromaeosaurids such as Achillobator giganticus (Perle et al. 1999) and Dromaeosaurus albertensis (Currie, Reference Currie1995).
The anteromedial process of the maxilla with a medial horizontal groove for contact with adjacent cranial elements in Ulughbegsaurus uzbekistanensis does not resemble the homologous feature in undisputed carcharodontosaurians (e.g. Neovenator salerii, Brusatte et al. Reference Brusatte, Benson and Hutt2008) but matches those in dromaeosaurids (Dromaeosaurus albertensis, Currie, Reference Currie1995; Utahraptor ostrommaysi, Fig. 3d).
Among the hundreds of non-avian theropod remains from the Bissekty Formation personally examined by AA and H-DS, there were no bones that could definitively be assigned to carcharodontosaurians. Tanaka et al. (Reference Tanaka, Anvarov, Zelenitsky, Ahmedshaev and Kobayashi2021) surmised that some of the larger isolated teeth assigned to Timurlengia euotica (Averianov & Sues, Reference Averianov and Sues2012) might belong to carcharodontosaurians. While this cannot be ruled out, at least none of the numerous teeth referred to Timurlengia euotica and examined by AA and H-DS shows features inconsistent with attribution to tyrannosauroids.
4. Conclusions
A pedal phalanx II-2 demonstrates the presence of a dromaeosaurid in the Bissekty Formation that attained larger body size than any other known member of this clade. Among dromaeosaurids, several taxa are distinguished by large body size: Utahraptor ostrommaysi with a femur length of 56.5 cm (Turner et al. Reference Turner, Makovicky and Norell2012); Austroraptor cabazai, from the Upper Cretaceous (Campanian–Maastrichtian) Allen Formation of Argentina, with a femur length of 56 cm (Novas et al. Reference Novas, Pol, Canale, Porfiri and Calvo2009); the possibly chimaeric Dakotaraptor steini, from the Upper Cretaceous (Maastrichtian) Hell Creek Formation of South Dakota, with a femur length of 55.8 cm (DePalma et al. Reference DePalma, Burnham, Martin, Larson and Bakker2015); and Achillobator giganticus with a femur length of 55 cm (Turner et al. Reference Turner, Makovicky and Norell2012). In addition, isolated teeth from the Lower Cretaceous (Barremian) Wessex Formation of England (Sweetman, Reference Sweetman2004) and the Upper Cretaceous (Campanian) Tar Heel Formation of North Carolina (Brownstein, Reference Brownstein2018) record dromaeosaurids as large as or larger than Deinonychus antirrhopus. The dimensions of the pedal phalanx II-2 of the giant Bissekty dromaeosaurid considerably exceed those of the corresponding phalanges in Achillobator giganticus and Austroraptor cabazai, respectively. Turner et al. (Reference Turner, Pol, Clarke, Erickson and Norell2007) estimated the total length of Achillobator giganticus at 4.85 m, and Novas et al. (Reference Novas, Pol, Canale, Porfiri and Calvo2009) provided an estimate of 5 m for the total length of Austroraptor cabazai. Achillobator giganticus and Utahraptor ostrommaysi are placed in Dromaeosaurinae (Turner et al. Reference Turner, Makovicky and Norell2012; Powers et al. Reference Powers, Fabbri, Doschak, Bhullar, Evans, Norell and Currie2022). By contrast, Austroraptor cabazai is a representative of Unenlagiinae, a predominantly South American clade of Dromaeosauridae. Thus, evolution toward very large body-size occurred at least twice among Dromaeosauridae (Wang et al. Reference Wang, Zhang, Tan, Jiangzuo, Zhang and Tan2022). Giant (>4 m long) dromaeosaurids clearly were apex predators in several Cretaceous terrestrial ecosystems.
The holotypic maxilla of Ulughbegsaurus uzbekistanensis presents several features that are found in both carcharodontosaurians and dromaeosaurids. In view of its highly fragmentary nature and the lack of unambiguous apomorphies linking it to any particular clade of non-avian theropods, the phylogenetic position of Ulughbegsaurus uzbekistanensis remains unresolved. It does not definitively establish the presence of carcharodontosaurian theropods in the Bissekty Formation. In the absence of autapomorphies or a diagnostic combination of character-states for Ulughbegsaurus uzbekistanensis, we also consider this binomen a nomen dubium.
Acknowledgements
H-DS thanks PJ Currie and AH Turner for sharing information on Achillobator giganticus, and AK Behrensmeyer for discussions concerning bone erosion. MJ Powers and CD Brownstein provided comments on the manuscript. DL Brinkman kindly provided photographs of the pedal phalanx II-2 of YPM VP.005205.



