



Non-technical Summary
There are very few articles dedicated to the study of crustacean remains in the marine sediments of the Mesozoic despite the widespread distribution of such remains in Central European Russia. This research is the first general study on decapods of the Ryazan region of Central European Russia and is focused on the Callovian of that area. The Ryazan region is renowned for its large number of outcrops and high concentration of well-preserved marine decapod fossils. Two genera and five species of crustaceans are identified for the first time from Central European Russia. In addition, diagnostic signs of previously noted taxa are factually discussed.
Introduction
European decapods from the Middle Jurassic are a highly diverse and abundant group of invertebrates (Schweitzer and Feldmann, Reference Schweitzer and Feldmann2014). The excellent preservation of representative samples of the rich decapod fauna is well known for Dendrobranchiata Bate, Reference Bate1888, Erymida Van Straelen, Reference Van Straelen1925, Glypheoidea von Zittel, Reference von Zittel1885, and other groups from the La Voulte Lagerstätte (Callovian of Central France) (Etter, Reference Etter, Bottjer, Etter, Hagadorn and Tang2002; Charbonnier et al., Reference Charbonnier, Vannier, Hantzpergue and Gaillard2010; Chény et al., Reference Chény, Charbonnier and Audo2023). Crônier and Courville (Reference Crônier and Courville2004) have described the local Bajocian–Oxfordian decapod fauna of erymids and pagurids from Etrochey, Northern France. Several recent studies were dedicated to crustaceans from the Kimmeridgian and Tithonian of Solnhofen, Germany, where the richest crustacean assemblage was discovered (Schweigert and Garassino, Reference Schweigert and Garassino2003, Reference Schweigert and Garassino2004; Garassino and Schweigert, Reference Garassino and Schweigert2006). Active investigations of erymid and glypheid lobsters significantly enriched the available evidence of crustacean diversity (Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013, Reference Charbonnier, Audo, Caze and Biot2014a; Schweigert, Reference Schweigert2013; Devillez and Charbonnier, Reference Devillez and Charbonnier2017, Reference Devillez and Charbonnier2019).
Whereas the crustacean assemblages of Western Europe were thoroughly studied, those of European Russia got little attention, despite the high abundance of the crustaceans discovered in the Middle and Late Jurassic in the region. There are only a few reliable publications (Trautschold, Reference Trautschold1866; Lahusen, Reference Lahusen1894; Gerasimov, Reference Gerasimov1955; Gerasimov et al., Reference Gerasimov, Mitta and Kochanova1995, Reference Gerasimov, Mitta, Kochanova and Tesakova1996) describing four species of Erymidae and Glypheidae. According to Gerasimov's (Reference Gerasimov1955) work, the only species, Eryma quadriverrucatum Trautschold, Reference Trautschold1866, is found in the middle–upper Callovian and Oxfordian of Central European Russia; the two species of Eryma (E. mosquensis Lahusen, Reference Lahusen1894 and E. gracilimanum Lahusen, Reference Lahusen1894), and the single glypheid taxon were spotted in the Tithonian (regional stage is the Volgian; Rogov, Reference Rogov, Rocha, Pais, Kullberg and Finney2014). However, by the latest revision of the erymids (Devillez and Charbonnier, Reference Devillez and Charbonnier2021), Eryma mosquensis Lahusen, Reference Lahusen1894 is considered to be a junior synonym for E. quadriverrucatum. Another Tithonian species, E. gracilimanum Lahusen, Reference Lahusen1894, is included in E. lerasi (Étallon, 1861). Finally, Glypheopsis vosinskyi Lahusen, Reference Lahusen1894 (Glypheidae), which is the third species mentioned by Gerasimov (Reference Gerasimov1955), was recently attributed to Erymidae (Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013; Devillez and Charbonnier, Reference Devillez and Charbonnier2021). In addition, several specimens of erymids and two unidentified species of Prosopidae were found in the late Tithonian of Moscow and the Moscow Region and described by Gerasimov (Reference Gerasimov1955). However, the present authors have doubts concerning the accuracy of the determination of those damaged remains. In general, the poor and incomplete preservation of crustaceans made it challenging to account for the accurate specifications of these samples. Since 1996, a few additional papers were published. Tesakova (Reference Tesakova2008) reported the presence of Mecochirus sp. in the Callovian of the Dubki locality (Saratov Region, Russia). In addition, Archeosoleonocera straeleni Carriol and Riou, Reference Carriol and Riou1991 was reported from Mikhailovtsement (Ryazan region, Russia) by Shmakov (Reference Shmakov2016). Unfortunately, both publications contained no pictures of the specimens; the specimen of Archeosolenocera from Shmakov (Reference Shmakov2016) is included in the present work. Later, Mironenko (Reference Mironenko2020) reported the occurrence of a hermit crab buried in an ammonite shell (Tithonian of Moscow). The present paper provides new and detailed information on several recently discovered samples of crustaceans that have never been previously reported for the area.
Geological setting
The material described herein was found in the Gorenka, Spartak, and Mikhailovtsement quarries (Mikhailov District, Ryazan Region) and Nikitino Lagerstätte (Spassk District, Ryazan region) (Fig. 1). The Jurassic deposits in all the sections include the Callovian and lower and middle Oxfordian. The reddish and brown sandstones and silts of the lower Callovian (Koenigii zone) contain mostly bivalves, belemnites, and rarer nuclei of ammonites, which are more abundant in Gorenka (Figs. 1, 2) and Spartak (Figs. 1, 3) locations. The marine deposits of Mikhailovtsement (Figs. 1, 4) were previously described (Kiselev and Rogov, Reference Kiselev and Rogov2018). The only report of Eryma is from the Mikhailovtsement. The middle Callovian (Jason zone) is composed of brown sands and red-brown ferruginous sandstones with carbonate concretions including a few ammonites. The Coronatum zone of the middle Callovian includes brown sands, light brownish-gray siltstone, and dark-brown clay. The upper Callovian (Phaeinum and Proniae subzones of the Athleta zone) is similar to the middle Callovian and consists of brownish-gray clay with a rich assemblage of invertebrate fossils that were frequently preserved as pyrite molds. These layers have a thickness of up to 8 m. Small ferruginous oolites are observed at the top of the Proniae subzone. The dense brownish-gray clay of the Lamberti zone contains small ferruginous oolites and larger yellowish-gray marlstone concretions with an assemblage of cephalopods, gastropods, bivalves, polychaetes, and rarer crinoids or marine reptiles. The layer of phosphatized marlstone oolites is found at the top of this zone with multiple decapod fossils and pyrite inclusions. The concretions are usually less than 10 cm in diameter, with a pale gray surface and black dense kernel. The crustaceans are preserved on the surface of oolites, partially submerged in the marlstone or inside it. The fossils are three-dimensional, occasionally deformed, black or rarely pale-colored, and often pyritized. The isolated pereopod and antennae segments are the most abundant, whereas the carapaces and chelae occur rarer, mostly as disarticulated remains.

Figure 1. Distribution of the studied localities. 1 = Gorenka quarry (54.238392, 38.848255); 2 = Spartak quarry (54.232961, 38.866298); 3 = Mikhailovtsement quarry (54.211668, 38.955771); 4 = Nikitino Lagerstätte (54.353765, 40.407202). The red stars mark the proper locations of the studied deposits.

Figure 2. The Callovian section of Gorenka quarry. The red star marks the layer where the crustacean remains were found.
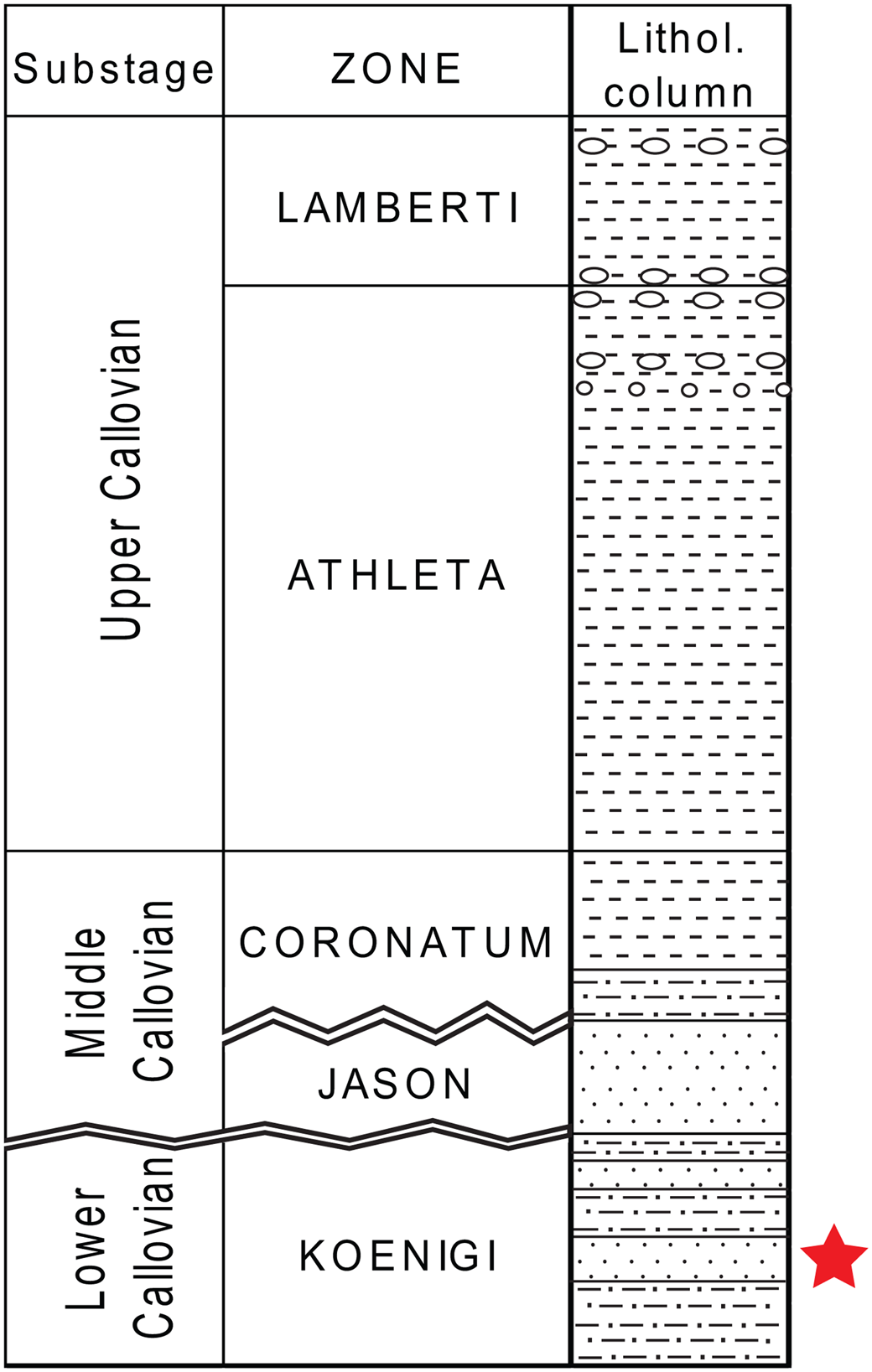
Figure 3. The Callovian section of Spartak quarry. The red star marks the layer where the crustacean remains were found.

Figure 4. The Callovian section of Mikhailovtsement quarry. The red star marks the layer where the crustacean remains were found.
The deposits of Nikitino (Figs. 1, 5) represent the Jurassic (Callovian and lower Oxfordian) and Lower Cretaceous. The shale marlstones of the Koenigi zone of the lower Callovian are reddish yellow with oolites. The Calloviense zone consists of various very argillaceous siltstones from reddish to gray in color. Both subzones are rich in invertebrates such as ammonites, bivalves, and gastropods, as well as remains of marine reptiles. The single occurrence of decapods from Nikitino is attributed to the Enodatum subzone. The Jason zone of the middle Callovian is composed of gray clays with large concretions at the base. The Coronatum zone consists of very argillaceous dark-brown siltstone with numerous tiny oolites and reddish-gray marlstone with a brownish-red surface. In the upper part of the section, beds of the upper Callovian dark-gray clays are exposed (Gulyaev, Reference Gulyaev2001).
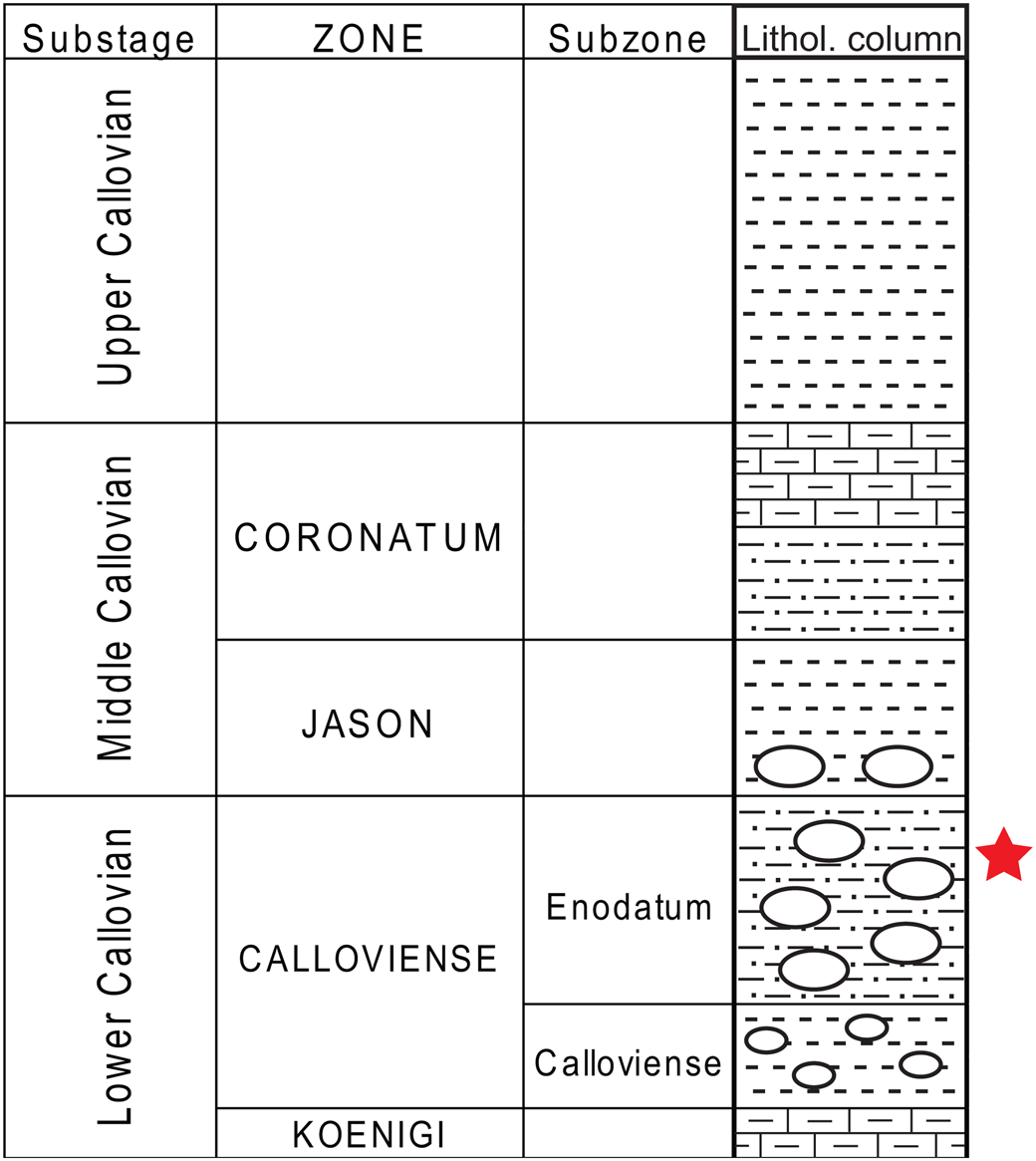
Figure 5. The Callovian section of Nikitino Lagerstätte. The red star marks the layer where the crustacean remains were found.
Materials and methods
Twenty-one specimens from the regional collections of the Paleontological Institute of Russian Academy of Science (PIN) and the Museum of Natural History at Moscow Children and Youth Centre for Ecology, Local History and Tourism (MCYCELHT) were used for the present research (Table 1). The crustacean remains were prepared using a pneumatic engraver. All the specimens were photographed with a gradual shift of focus by a digital camera Nikon D800; focusing was accomplished manually. The resulting series of photos were uploaded to the Helicon Focus Pro 6.0.18 software, and the processing of images was completed by the Adobe Photoshop CS2. The glaring specimens were sprayed with a powder of dry ammonium chloride to reduce surface gloss and to reveal their structure before taking pictures. All of the pictures and line drawings were made by I.A. Dadykin.
Table 1. List of the studied Callovian decapods of the Ryazan region.

The following abbreviations are used in the figures and text body: a = branchiocardiac groove; ar = antennal ridge; b = antennal groove; b1 = hepatic groove; bas = basipodite of uropod; c = postcervical groove; cd = cardiac groove; cp = carpus of P1; d = gastro-orbital groove; dac = dactylus of P1 chela; db = dactylar bulge; di = diaeresis; e1e = cervical groove; en = endopod of uropod; ex = exopod of uropod; i = inferior groove; ic = intercervical groove; in = index of P1 chela; ip = intercalated plate; or = orbital ridge; p = pereopods; P1 = pereopod 1; P5 = pereopod 5; pl = pleopods; pn = pleon; PoA = postorbital area of the cephalic region; pr = propodus of P1; r = rostrum; s = pleonal somite; sc = scaphocerite; ts = telson; u = uropods; χ = attachment site of the testis muscle; ω = attachment site of the mandibular muscle (for the explanatory diagrams, see Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013; Devillez and Charbonnier, Reference Devillez and Charbonnier2019, Reference Devillez and Charbonnier2021).
Repositories and institutional abbreviations
MCYCELHT = Museum of Natural History at Moscow Children and Youth Centre for Ecology, Local History and Tourism, Moscow, Russia; MNHN = Muséum national d'Histoire naturelle, Paris, France; NHMUK = Natural History Museum, London, UK; OSUG UJF-ID = Observatoire des Sciences de l'Univers de Grenoble, Institut Dolomieu, Grenoble, France; PIN = Paleontological Institute of Russian Academy of Science, Moscow, Russia; SMNS = Staatliches Museum für Naturkunde, Stuttgart, Germany.
Systematic paleontology
Order Decapoda Latreille, Reference Latreille1802
Infraorder Dendrobranchiata Bate, Reference Bate1888
Superfamily Penaeoidea Rafinesque, Reference Rafinesque1815
Penaeoidea gen. indet. sp. indet.
Figure 6.1–6.3
Description
The specimen PIN 5477/3771 exposes the isolated pleon with preserved P5 coxae. The pleonal somites are not carinate; each somite overlaps the next one; the pleurae are smooth and rounded. The coxae of P5 are globose. The pleopods are poorly preserved, biflagellate; flagellae are multiarticulate. The uropods are poorly preserved as well, ornamented by minute tubercles.

Figure 6. (1–3) Penaeoidea gen. indet. sp. indet. from Mikhailovtsement, upper Callovian (PIN 5477/3771): (1, 2) right lateral view; (3) uropods, ventral view. (4–7) Archeosolenocera sp. from Mikhailovtsement, upper Callovian (PIN 5477/2933): (4, 5) left lateral view; (6, 7) dorsal view. b = antennal groove; b1 = hepatic groove; di = diaeresis; e1e = cervical groove; en = endopod of uropod; ex = exopod of uropod; i = inferior groove; P5 = pereopod 5; pl = pleopods; pn = pleon; ts = telson; u = uropods. Scale bars = 10 mm.
Material
PIN 5477/3771 from the upper Callovian of Mikhailovtsement.
Remarks
The specimen PIN 5477/3771 is insufficiently preserved and cannot be attributed to any family with confidence. However, we attribute this specimen to Penaeoidea since it has the first pleonal somite overlapping the second one and biflagellate pleopods with multiarticulate flagellae (Garassino et al., Reference Garassino, Pasini and Vega2013). Another specimen (PIN 5477/2933) might be attributed to family Solenoceridae Wood-Mason in Wood-Mason and Alcock, Reference Wood-Mason and Alcock1891 on the basis of the cervical groove not reaching the dorsal margin, the antennal groove widening anteriorly, the subhorizontal hepatic groove, and the inferior groove directed frontward (Jauvion et al., Reference Jauvion, Charbonnier and Bernard2017). Unfortunately, the anterior part of the carapace is strongly damaged; thus, the presence of suprarostral spines characteristic of Solenoceridae (Jauvion et al., Reference Jauvion, Charbonnier and Bernard2017; Alencar et al., Reference Alencar, Pinheiro, Saraiva, Oliveira and Santana2018) could not be confirmed.
Family Solenoceridae Wood-Mason in Wood-Mason and Alcock, Reference Wood-Mason and Alcock1891
Genus Archeosolenocera Carriol and Riou, Reference Carriol and Riou1991
Type species
Archeosolenocera straeleni Carriol and Riou, Reference Carriol and Riou1991 by monotypy.
Archeosolenocera sp.
Figure 6.4–6.7
Description
The specimen PIN 5477/2933 includes a carapace and four pleonal segments. The carapace is elongate (74 mm long), laterally compressed, almost smooth. Its dorsal margin is straight; ventral margin is slightly convex. The rostrum is poorly preserved, wide. Cervical groove is deep and short, inclined, joined ventrally to straight antennal groove; antennal groove short, widening anteriorly; hepatic groove is elongate, subhorizontal, slightly inclined posteriorly, joined anteriorly to cervical and antennal grooves, forming an angle of 100° with the cervical groove; no spine at junction of the cervical and the hepatic grooves; inferior groove wide, inclined, joined to hepatic groove anteriorly to its junction to cervical groove. The pleon is laterally compressed; the pleonal segments are smooth, and the pleura are rounded.
Material
PIN 5477/2933 from the upper Callovian of Mikhailovtsement.
Remarks
Family Solenoceridae comprises three extinct monotypic genera (Alencar et al., Reference Alencar, Pinheiro, Saraiva, Oliveira and Santana2018): Archeosolenocera Carriol and Riou, Reference Carriol and Riou1991 (Middle Jurassic, France), Eogordonella Garassino et al., Reference Garassino, Bahrami, Yazdi and Vega2014 (Eocene, Iran), and Priorhyncha Alencar et al., Reference Alencar, Pinheiro, Saraiva, Oliveira and Santana2018 (Early Cretaceous, Brazil). Priorhyncha differs from our specimen in having a cervical groove only. Eogordonella has a serrate dorsal margin of the cephalic region and the hepatic, branchiocardiac, and submarginal carinae, which are absent in specimen PIN 5477/2933. We may attribute this specimen to Archeosolenocera due to its specific groove pattern: antennal, cervical, hepatic, and inferior grooves present; cervical groove well developed and not reaching the dorsal margin of the carapace; hepatic and antennal grooves represented; inferior groove slightly incurved and subparallel to the dorsal margin. However, the specimen PIN 5477/2933 differs from the diagnosis of Jauvion et al. (Reference Jauvion, Charbonnier and Bernard2017) in the absence of a hepatic spine and junction of inferior groove to hepatic groove at the level of the antennal groove, anteriorly to the junction of the cervical groove. Thus, PIN 5477/2933 probably does not belong to the single known species of the genus, A. straeleni Carriol and Riou, Reference Carriol and Riou1991 from the Callovian of France. Unfortunately, the preservation of our material is not quite good enough to clarify the taxonomy of the specimen. Therefore, we attribute the specimen PIN 5477/2933 to Archeosolenocera sp.
Infraorder Pleocyemata Burkenroad, Reference Burkenroad1963
Superfamily Glypheoidea von Zittel, Reference von Zittel1885
Family Mecochiridae Van Straelen, Reference Van Straelen1925
Genus Eumorphia von Meyer, Reference von Meyer1847
Type species
Eumorphia socialis (von Meyer, Reference von Meyer1841) (=Mecochirus socialis (von Meyer, Reference von Meyer1841)), by monotypy, from the upper Callovian of St. Ives, Huntingdonshire, Great Britain.
Description
All the specimens share similar morphological features and expose the carapaces with partially preserved thoracic appendages and pleon.

Figure 7. Eumorphia sp. (Mecochiridae) from Gorenka (upper Callovian), dusted with ammonium chloride: (1, 2) complete shrimp (PIN 5477/3788), left lateral view; (3, 4) complete shrimp (PIN 5477/4547), left lateral view; (5–8) complete shrimp (PIN 5477/3787): (5, 6) left lateral view; (7, 8) pleon, dorsal view. a = branchiocardiac groove; b1 = hepatic groove; c = postcervical groove; cd = cardiac groove; cp = carpus of P1; e1e = cervical groove; en = endopod of uropod; ex = exopod of uropod; or = orbital ridge; p = pereopods; P1 = pereopod 1; pn = pleon; pr = propodus of P1; ts = telson. Scale bar = 10 mm.

Figure 8. Eumorphia sp. (Mecochiridae) from Gorenka (upper Callovian), dusted with ammonium chloride: (1–3) complete shrimp (PIN 5477/3789): (1) left lateral view, (2, 3) carapace, dorsal view; (4–6) complete shrimp (PIN 5477/4549): (4, 5) right lateral view, (6) left lateral view; (7, 8) complete shrimp (PIN 5477/4548): (7) left lateral view, (8) right lateral view. a = branchiocardiac groove; b1 = hepatic groove; bas = basipodite of uropod; c = postcervical groove; cd = cardiac groove; e1e = cervical groove; en = endopod of uropod; ex = exopod of uropod; or = orbital ridge; p = pereopods; pn = pleon; r = rostrum; ts = telson. Scale bar = 10 mm.
Carapace.—The carapace is ovoid in the lateral view; it is laterally compressed (carapace length is ~10–15 mm; carapace height is ~7 mm). The rostrum is short, pointed, smooth, with medial dorsal longitudinal depression. The antennal–pterygostomial margin is slightly convex. The cervical groove is long, deep, oblique, joining to the dorsal margin at an angle of ~37°, delimiting the short cephalic region. The cephalic region is almost smooth, bearing a single orbital ridge. The postcervical and branchiocardiac grooves are subparallel, shallow, reaching the posterior margin of the carapace, forming an angle of ~18° with the dorsal margin. The narrow lobe between the postcervical and the branchiocardiac grooves is represented. The postcervical groove is narrow and incurved anteriorly, slightly curved medially, joined dorsally to the cardiac groove; the branchiocardiac groove is narrow and straight, joined ventrally to the hepatic groove. The cardiac groove is elongate, straight or slightly incurved backward, inclined forward, and reaching the dorsal margin at an angle of ~60°; the cardiac grooves do not join at the midline of the carapace. The hepatic groove is shallow and deepening posteriorly. Its posterior end is turning forward, forming a circular slightly convex hepatic lobe. The cephalic, the hepatic, the pterygostomial, and the cardiac regions are almost smooth, ornamented only by a few minute tubercles. The branchial region is densely tuberculate, with the largest tubercules situated at the center of the region.
Pleon.—The pleon is about twice as long as the carapace. The pleomeres are bearing longitudinal carina laterally, with rounded dorsal margin almost smooth. The pleurae are rounded and tuberculate, bearing short transverse grooves anteriorly and posteriorly. The s1 is short; s2 is the longest one; s3–5 are of similar length; s6 is short and subtriangular. The telson is subrectangular, flattened, smooth, and posteriorly rounded.
Thoracic appendages.—The P1–5 are strongly damaged; subchelate P1, slender, 1.5 times longer than the carapace; P1 carpus flattened laterally, with a row of tubercles at posterior edge; P1 propodus elongate, subtriangular in section, densely tuberculated; P2–5 thin, slightly flattened laterally, almost smooth.
Pleonal appendages.—The pleopods are not preserved. The uropods are biramous; the basipodite is smooth and globose; the exopod and the endopod are of similar shape, ovoid, with a wide longitudinal carina narrowing distally, ornamented by sparse tubercles; the exopod lacks diaeresis; outer margin of exopod is smooth.
Material
PIN 5477/3787, PIN 5477/3788, PIN 5477/3789, PIN 5477/4547, PIN 5477/4548, and PIN 5477/4549 from the upper Callovian of Gorenka.
Remarks
All specimens discussed in the preceding (Figs. 7, 8) are assigned to Mecochiridae on the basis of the following characteristic features (after Karasawa et al., Reference Karasawa, Schweitzer and Feldmann2013): the presence of the cervical, postcervical, and branchiocardiac grooves; the low angles of the cervical and the branchiocardiac grooves to dorsal surface; the closely spaced and subparallel postcervical and branchiocardiac grooves; the absence of an inferior groove; the elongated pereopod 1; and the presence of cephalic ridges (Karasawa et al., Reference Karasawa, Schweitzer and Feldmann2013).
Family Mecochiridae remains unrevised, and its systematics is still in progress (S. Charbonnier, personal communication, 2023). To date, mecochirids include four valid genera: Atherfieldastacus Robin et al., Reference Robin, Charbonnier, Merle, Simpson, Petit and Fernandez2016, Huhatanka Feldmann and West, Reference Feldmann and West1978, Meyeria M'Coy, Reference M'Coy1849, and Mecochirus Germar, Reference Germar and Keferstein1827. Two more genera are considered to be invalid: Jabaloya Garassino, Artal, and Pasini, 2009 is a junior synonym for Meyeria, and Meyerella Simpson and Middleton, Reference Simpson and Middleton1985 is recently replaced by Atherfieldastacus (Robin et al., Reference Robin, Charbonnier, Merle, Simpson, Petit and Fernandez2016). A relation of Pehuenchia Rusconi, Reference Rusconi1948 and Zapalianassa Schweitzer, Feldmann, and Casadio, 2022 (Late Jurassic, Argentina) to Mecochiridae was recently argued (Schweitzer et al., Reference Schweitzer, Feldmann and Casadío2022). Family Mecochiridae includes both Jurassic and Cretaceous species (Glaessner, Reference Glaessner and Moore1969; Simpson and Middleton, Reference Simpson and Middleton1985; De Grave et al., Reference De Grave, Pentcheff, Ahyong, Chan and Crandall2009; Garassino et al., Reference Garassino, Artal and Pasini2009; Astrop, Reference Astrop2011; Karasawa et al., Reference Karasawa, Schweitzer and Feldmann2013; Robin et al., Reference Robin, Charbonnier, Merle, Simpson, Petit and Fernandez2016; Schweitzer et al., Reference Schweitzer, Feldmann and Casadío2022).
The diagnostic features of studied specimens are as follows: a single cephalic ridge; subparallel postcervical and branchiocardiac grooves reaching the posterior margin of the carapace; the cardiac groove represented; the inferior groove absent; the hepatic groove strongly incurved, forming a hepatic lobe; the hepatic ridge absent; no diaeresis on the exopod of uropods, the latter one with smooth outer margin (Figs. 7, 8). On the basis of these characteristics, discussed specimens do not belong to Meyeria (three cephalic carinae and the inferior groove is represented), Mecochirus (three cephalic carinae, the cardiac groove is absent, the exopod with a diaeresis), or Atherfieldastacus (three cephalic carinae, the exopod with a diaeresis and serrate outer margin) (Garassino et al., Reference Garassino, Artal and Pasini2009; Robin et al., Reference Robin, Charbonnier, Merle, Simpson, Petit and Fernandez2016; Charbonnier et al., Reference Charbonnier, Audo, Garassino, Simpson, Gèze and Azar2021). S. Charbonnier suggested that the name Eumorphia von Meyer, Reference von Meyer1847 continue to be considered a junior synonym of Mecochirus for individuals without the diaeresis and with the cardiac groove represented (S. Charbonnier, personal communication, Reference Chény, Charbonnier and Audo2023). Thus, we propose the name Eumorphia sp. for all of the studied material in this section. The record of Mecochirus sp. by Tesakova (Reference Tesakova2008) from the upper Callovian of Dubki quarry, Saratov region, probably relates to that taxon. (For more information on Eumorphia socialis (von Meyer, Reference von Meyer1841), see Carter, Reference Carter1886; Woods, Reference Woods1928; Glaessner, Reference Glaessner and Moore1969).
Family Glypheidae Winkler, Reference Winkler1882
Glypheidae gen. indet. sp. indet.
Figure 9.1, 9.2
Description
The specimen PIN 5477/3724 exposes the isolated posterior part of the carapace. The carapace is elliptical in dorsal view (length of preserved part is ~20 mm); the postcervical, branchiocardiac, and cardiac grooves are represented; the postcervical and branchiocardiac grooves converge dorsally, forming an elongate lobe; the postcervical groove is shallow; the branchiocardiac groove is deep and wide, reaching the dorsal margin of the carapace at an angle of ~20°; the branchiocardiac grooves are connected dorsally; the cardiac groove is shallow and short, inclined forward; the carapace is ornamented by large pits.

Figure 9. Glypheidae from Middle Jurassic of Ryazan region: (1, 2) Glypheidae gen. indet. sp. indet. from Nikitino, middle Callovian (PIN 5477/3724): (1) dorsal view, (2) right lateral view; (3, 4) Glypheopsis sp. from Mikhailovtsement, upper Callovian: (3) carapace (PIN 5477/3786), left lateral view, (4) carapace (PIN 5477/4542, dusted with ammonium chloride), left lateral view; (5, 6) Glypheopsis aff. G. etalloni (Oppel, Reference Oppel1861): carapace from Gorenka, upper Callovian (PIN 5477/4536, dusted with ammonium chloride), right lateral view. a = branchiocardiac groove; b = antennal groove; b1 = hepatic groove; c = postcervical groove; cd = cardiac groove; e1e = cervical groove; i = inferior groove, ic = intercervical groove. Scale bars = 10 mm.
Material
PIN 5477/3724 from the early Callovian of Nikitino.
Remarks
The family has numerous paleontological records for the Mesozoic and Cenozoic (Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013, 2014a). To date, eight extinct and two extant genera of Glypheidae are known (Charbonnier et al., Reference Charbonnier, Audo, Barriel, Garassino, Schweigert and Simpson2014a). We attribute the specimen PIN 5477/3724 to the family Glypheidae because of the following taxonomically important features: the postcervical and branchiocardiac grooves are subparallel and converging dorsally, forming an elongate lobe. The presence of the cardiac groove, a single lobe between the postcervical and branchiocardiac grooves, and the absence of the laterocardiac groove are diagnostic for both Glypheopsis Beurlen, Reference Beurlen1928 and Trachysoma Bell, Reference Bell1858. Unfortunately, the condition of the discussed specimen does not allow further determination.
Genus Glypheopsis Beurlen, Reference Beurlen1928
Type species
Orphnea ornata Quenstedt, 1857 by the subsequent designation of Glaessner (Reference Glaessner and Pompeckj1929) from the Callovian of Pfullingen, Baden-Württemberg, Germany.
Glypheopsis sp.
Figure 9.3, 9.4
Description
The specimens PIN 5477/3786 and PIN 5477/4536 expose the posterior parts of carapaces. The cervical groove is deep, forming an angle of ~70° with the dorsal margin, joining ventrally to the antennal groove, with delimiting elongate cephalic region (it is absent on specimen PIN 5477/3786). The cardiac groove is shallow and almost straight, inclined anteriorly, and forming an angle of ~47° with the dorsal margin. The hepatic groove is deep and bilobed, joined to the cervical groove; the adductor muscle area is not inflated. The intercervical groove is weak, slightly incurved upward, delimiting small hepatic lobe. The branchial, pterygostomial, and hepatic regions are densely ornamented by tubercles; those of the branchial region decrease in size ventrally; the cardiac region with scarce and largest tubercles.
Material
PIN 5477/3786 and PIN 5477/4542 from the upper Callovian of Mikhailovtsement.
Remarks
The discussed specimens might be attributed to Glypheopsis Beurlen, Reference Beurlen1928 on the basis of the postcervical and branchiocardiac grooves forming a single elongate lobe; the cardiac groove represented; the laterocardiac groove absent; the hepatic groove bilobed; the intercervical groove represented; the gastro-orbital groove absent (Charbonnier et al., Reference Charbonnier, Audo, Barriel, Garassino, Schweigert and Simpson2014a). To date, the 26 species of Glypheopsis are reported from the Early Jurassic to the Eocene (Garassino et al., Reference Garassino, Charbonnier, Nyborg and Fam2019; Becker et al., Reference Becker, Fraaije and Mulder2021). Among them, the seven species occur in the Callovian and Oxfordian of Europe (Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013).
The ornamentation pattern of specimens PIN 5477/3786 and PIN 5477/4542 is characteristic for Glypheopsis etalloni (Oppel, Reference Oppel1861) and G. pseudomuensteri (Martin, Reference Martin1961) (both from the Oxfordian of France) or G. calloviensis (Woods, Reference Woods1927) (late Callovian of the United Kingdom) (Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013). Here we identify the specimens PIN 5477/3786 and PIN 5477/4542 only at the generic level.
Glypheopsis aff. G. etalloni (Oppel, Reference Oppel1861)
Figure 9.5, 9.6
Type material
Lectotype MNHN.F.A29575, collection of Étallon, Muséum national d'Histoire naturelle, Paris, France (Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013, p. 122, fig. 237).
Description
The specimen PIN 5477/4536 is a well-preserved carapace with fragments of P1. The cervical groove is deep. The carapace is elongate in the lateral view (length of carapace is ~34 mm; height of carapace is ~15 mm). The rostrum is not preserved. The cervical groove is deep, forming an angle of ~70° with the dorsal margin, joining ventrally to the antennal groove, with delimiting elongate cephalic region. The cephalic region bears four longitudinal spiny carinae, which do not reach the cervical groove: the orbital, the gastro-orbital, the antennal, and the supra-antennal carinae. The intercarinal spaces are smooth; the antennal region is ornamented by dense tubercles. The antennal groove is narrow, deep, strongly incurved posteriorly, and slightly incurved upward at its middle part. The postcervical and branchiocardiac grooves converge ventrally and dorsally and form a single elongate lobe; the postcervical groove is slightly tortuous, converging with the hepatic groove ventrally, connected to the intercervical and cardiac grooves. The branchiocardiac groove is almost straight, bending up posteriorly and reaching the dorsal margin of the carapace at an angle of ~75°. The P1 propodus (partially preserved) is ornamented by longitudinal rows of spines.
Material
PIN 5477/4536 from the upper Callovian of Gorenka.
Remarks
The specimen PIN 5477/4536 (Fig. 9.5, 9.6) shares the diagnostic features of Glypheopsis etalloni: four longitudinal spiny carinae on the cephalic region; weakly inflated adductor muscle area; the ornamentation by tubercles decreasing in size ventrally in the branchial region (Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013, figs. 237–246). However, the specimen PIN 5477/4536 is distinguished from G. etalloni by the presence of the bilobed hepatic groove (sinuous in G. etalloni) and the flat adductor muscle area (inflated in G. etalloni); in addition, the space between the antennal and supra-antennal carinae is smooth in specimen PIN 5477/4536 (it bears a row of tubercles in G. etalloni). G. pseudomuensteri (Martin, Reference Martin1961) from the Oxfordian of France is a very similar species to G. etalloni and may be its junior synonym (Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013). The specimen PIN 5477/4536 differs from this species by having four cephalic carinae (three for G. pseudomuensteri), tuberculate pterygostomial region (smooth for G. pseudomuensteri), and flat adductor muscle insertion area (Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013, figs. 283, 284). The specimen PIN 5477/4536 is distinct from other same-aged Glypheopsis species by having four cephalic carinae (five for G. calloviensis (Woods, Reference Woods1927), three for G. ornata (Quenstedt, Reference Quenstedt1858), G. bronnii (Roemer, Reference Roemer1839), G. trouvillensis Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013, and G. voultensis Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013), the ornamentation of the carapace by tubercles decreasing in size ventrally (uniform tubercles for G. ornata, pits for G. bronnii and G. trouvillensis, and almost absent for G. voultensis), and flat adductor muscle insertion area (inflated for G. ornata, G. calloviensis, G. bronnii, and G. voultensis). A biconvex hepatic groove is observed in many Cretaceous and Cenozoic Glypheopsis species (Charbonnier et al., Reference Charbonnier, Garassino, Schweigert and Simpson2013). However, no such taxa have been reported from the Callovian to Oxfordian. Hence, we propose a preliminary combination Glypheopsis aff. G. etalloni for the specimen PIN 5477/4536.
Infraorder Astacidea Latreille, Reference Latreille1802
Superfamily Erymoidea Van Straelen, Reference Van Straelen1925
Family Erymidae Van Straelen, Reference Trautschold1925
Genus Eryma von Meyer, Reference von Meyer1840b
Type species
Macrourites modestiformis von Schlotheim, Reference von Schlotheim1822 by subsequent designation of Glaessner, Reference Glaessner and Pompeckj1929, from the Kimmeridgian–Tithonian of Eichstätt, Bavaria, Germany.
Material
PIN 5477/0925, PIN 5477/3784, and MCYCELHT 2/226 from the upper Callovian of Mikhailovtsement; PIN 5477/3765 from the lower Callovian of Spartak; PIN 5477/4535, PIN 5477/4537, PIN 5477/4538, PIN 5477/4539, and PIN 5477/4953 from the upper Callovian of Gorenka.
Eryma aff. E. ventrosum (von Meyer, Reference von Meyer1835)
Figure 10.1–10.3
Type material
Probably lost, cast MNHN.F.B12484 (Devillez and Charbonnier, Reference Devillez and Charbonnier2021, fig. 11A, B).

Figure 10. (1–3) Eryma aff. E. ventrosum (von Meyer, Reference von Meyer1835): (1, 2) carapace from Mikhailovtsement, upper Callovian (PIN 5477/0925), left lateral view (dusted with ammonium chloride), (3) carapace from Gorenka, upper Callovian (PIN 5477/4535, dusted with ammonium chloride), left lateral view; (4) Eryma ornatum (Quenstedt, Reference Quenstedt1858), P1 chela from Spartak, lower Callovian (PIN 5477/3765), dorsal view. a = branchiocardiac groove; ar = antennal ridge; b = antennal groove; b1 = hepatic groove; c = postcervical groove; cp = carpus of P1; d = gastro-orbital groove; dac = dactylus of P1 chela; e1e = cervical groove; i = inferior groove; in = index of P1 chela; or = orbital ridge; p = pereopods; PoA = postorbital area of the cephalic region; pr = propodus of P1; r = rostrum; sc = scaphocerite. Scale bars = 10 mm.
Description
Carapace.—The carapace of PIN 5477/0925 is ovoid in the lateral view (carapace length is ~56 mm; carapace height is ~26 mm). The rostrum is short, pointed; the dorsal margin of the rostrum is serrate. The postorbital area is represented. The cervical groove is strongly inclined dorsally and slightly inclined ventrally, joined to the antennal groove ventrally, delimiting short cephalic region. Two cephalic carinae, converging toward the posterior, reach the middle part of the cephalic region; both carinae bear spines. The orbital spine is represented. The gastro-orbital groove is very short. The antennal groove is wide, shallow, slightly convex posteriorly. The postcervical groove is deep, wide, joining to the branchiocardiac groove at the carapace mid-height; the branchiocardiac groove is deep, shallower, slightly incurved at the junction with the postcervical groove, joins ventrally to the hepatic and inferior grooves.
The length of the carapace PIN5477/4535 is ~49 mm; the height is ~23 mm. The specimen shares features of PIN 5477/0925, despite the surface of the pterygostomial, hepatic, cardiac, and branchial regions being strongly damaged. The hepatic groove is concavo-convex, joining anteriorly to the cervical groove; the χ and ω areas are strongly damaged. The inferior groove is represented on the specimen PIN 5477/4535, deep and wide, incurved forward.
Carapace ornamentation.—The cephalic region bears two to three large spines between the orbital and antennal rows; the middle part of the cephalic region is approximately smooth, with circular depressions only. The gastric, cardiac, and branchial regions of PIN 5477/0295 are ornamented by sparse tubercles interspersed with circular depressions; the antennal region is ornamented by large tubercles without depressions.
Cephalic appendages.—Antennae are partially preserved in PIN 5477/0925. The scaphocerite is ~8 mm long, trapezoidal, with an elongated and slightly narrowing dorsal expansion equal in length to the basal part of the segment; the scaphocerite is smooth. Flagellum is strongly damaged.
Thoracic appendages.—The appendages are represented on the specimen PIN 5477/0925. The propodus is ~18 mm long, compressed dorsoventrally, ornamented by multiple tubercles; the pereopods are smooth.
Material
PIN 5477/0925 from the upper Callovian of Mikhailovtsement; PIN 5477/4535 from the upper Callovian of Gorenka.
Remarks
The specimens PIN 5477/0925 and PIN 5477/4535 belong to Erymidae on the basis of the cervical, postcervical, hepatic, and inferior grooves present; the short and shallow gastro-orbital groove; and the delimited postorbital area. I addition, the P1 propodus is preserved on PIN 5477/0925 (Fig. 10.1, 10.2). For a diagnosis of the family, see Devillez et al. (Reference Devillez, Charbonnier and Pezy2018).
Both specimens share features of Eryma von Meyer, 1840 (von Meyer, Reference von Meyer1840a): the postcervical groove ventrally joins to the branchiocardiac groove; the branchiocardiac groove ventrally joins to the hepatic groove; the hepatic groove is concavo-convex, forming χ and ω areas; the orbital spine of the orbital row is represented. (For a diagnosis of Eryma, see Devillez et al., Reference Devillez, Charbonnier and Pezy2018; Devillez and Charbonnier, Reference Devillez and Charbonnier2021).
The specimens are similar to E. ventrosum (von Meyer, Reference von Meyer1835) from the Late Jurassic of Europe in having the junction of the postcervical and branchiocardiac grooves at the mid-height of the carapace, as well as having the same cervical, hepatic, gastro-orbital, and inferior groove patterns. However, the studied material differs from typical E. ventrosum (Devillez and Charbonnier, Reference Devillez and Charbonnier2021, figs. 4, 5) by the presence of a few large spines at the space between the cephalic rows (absent for E. ventrosum); serrate rostrum (smooth for E. ventrosum); ornamentation by tubercles interspersed with circular depressions or without pits (tubercles with crescent-shaped depressions for E. ventrosum). Thus, we propose a preliminary name Eryma aff. E. ventrosum (von Meyer, 1835) for the studied specimens.
Eryma ornatum (Quenstedt, Reference Quenstedt1858)
Figure 10.4
Type material
GPIT/CU/00349, collection of Fachbereichs Geowissenschafte, Eberhard Karls Universität, Tübingen, Germany (Devillez and Charbonnier, Reference Devillez and Charbonnier2019, fig. 8A).
Description
Isolated P1 chela. The propodus is short (propodus length without index ~13 mm; width ~9 mm), trapezoidal, laterally compressed; the inner and outer margins are narrow; the ventral surface of the propodus with a longitudinal inflexion. The dactylar bulge is narrow, delimited by a deep groove. The fingers are thin (length ~25 mm), slightly incurved, progressively narrowing, and equal in length; index is wide at its base, with a steep incurvation and a wide shallow longitudinal groove on ventral surface. The occlusal margins have small, closely spaced teeth. The ornamentation of the chela is represented by small, dense tubercles with crescent-shaped depressions.
Material
PIN 5477/3765 from the lower Callovian of Spartak.
Remarks
The specimen PIN 5477/3765 is attributed to family Erymidae and genus Eryma on the basis of the following: P1 is chelate; P1 propodus is laterally compressed, with narrow inner and outer margins; the dactylar bulge is narrow, delimited by a groove; the P1 fingers are equal in length and thin; they are longer than the propodus; index wider than the dactylus.
The specimen PIN 5477/3765 can be attributed to E. ornatum (Quenstedt, Reference Quenstedt1858) from the Middle Jurassic of Europe because the propodus is trapezoidal, the fingers are slightly incurved, the index base does not form an angle with its distal part, and the ornamentation of P1 is represented by tubercles with crescent-shaped depressions.
The closely related species Eryma compressum (Eudes-Deslongchamps, Reference Eudes-Deslongchamps1842) differs from E. ornatum by having a wider base of the index forming a slight angle between proximal and distal parts of the index (J. Devillez, personal communication, 2021; Devillez and Charbonnier, Reference Devillez and Charbonnier2019, fig. 5D, F). The specimen PIN 5477/3765 lacks this feature; no curvature is observed in a proximal part of the index (Fig. 10.4). The specimen differs from E. mandelslohi (von Meyer, Reference von Meyer1840b) (Callovian, Europe) by having a trapezoidal propodus and slightly incurved dactylus (Devillez and Charbonnier, Reference Devillez and Charbonnier2019, fig. 7E, F); from E. ventrosum (Callovian to Kimmeridgian, Europe) and Eryma lerasi (Étallon, Reference Étallon1861) (Late Jurassic, Europe and European Russia) by possessing a trapezoidal propodus (Devillez and Charbonnier, Reference Devillez and Charbonnier2019, Reference Devillez and Charbonnier2021); from Eryma georgeii Carter, Reference Carter1886, (Oxfordian, United Kingdom) by fine ornamentation of chela (Devillez and Charbonnier, Reference Devillez and Charbonnier2019). Therefore, we suggest attributing the specimen PIN 5477/3765 to E. ornatum.
Eryma quadriverrucatum Trautschold, Reference Trautschold1866
Figure 11
Type material
NHMUK 22917, collection of the Natural History Museum, London, UK (Devillez and Charbonnier, Reference Devillez and Charbonnier2019, p. 10, fig. 5A, B).

Figure 11. Eryma quadriverrucatum Trautschold, Reference Trautschold1866: (1) well-preserved specimen from Mikhailovtsement, upper Callovian (MCYCELHT 2/226, dusted with ammonium chloride), left lateral view; (2) carapace from Gorenka, upper Callovian (PIN 5477/4537, dusted with ammonium chloride), right lateral view; (3, 4) carapace from Mikhailovtsement, upper Callovian, left lateral view (PIN 5477/3784). a = branchiocardiac groove; b = antennal groove; b1 = hepatic groove; c = postcervical groove; d = gastro-orbital groove; e1e = cervical groove; i = inferior groove; ip = intercalated plate; or = orbital ridge; pn = pleon; PoA = postorbital area of the cephalic region; r = rostrum; χ = attachment site of the testis muscle; ω = attachment site of the mandibular muscle. Scale bars = 10 mm.
Description
MCYCELHT 2/226 exposes a well-preserved carapace with joined pleon and thoracic appendages (Fig. 11.1); PIN 5477/4537 (Fig. 11.2) and PIN 5477/3784 (Fig. 11.3, 11.4) expose the isolated carapaces.
Carapace.—The carapace is elongate in the lateral view and subcylindrical in the cross section (length of specimen MCYCELHT 2/226 ~38 mm; height ~18 mm). The rostrum of specimen PIN 5477/3784 (Fig. 11.3, 11.4) is short and spineless. The postorbital area is narrow; the intercalated plate is fusiform (represented on specimen PIN 5477/3784). The cervical groove is strongly inclined dorsally, slightly inclined ventrally, joins to antennal groove ventrally, delimiting short cephalic region. Two cephalic carinae are represented, converging toward the middle part of the cephalic region; the orbital spine is represented. The antennal groove is deep and short, slightly convex posteriorly. The gastro-orbital groove is very short. The postcervical and branchiocardiac grooves are deep and subparallel, not reaching the dorsal margin of the carapace; the postcervical groove is inclined, slightly incurved, joins to the branchiocardiac groove at the carapace mid-height, bears a short ventral extension; the branchiocardiac groove is inclined, strongly incurved at its junction to the postcervical groove, joining ventrally to the hepatic groove. The hepatic groove is shallow and concavo-convex, joining anteriorly to the cervical groove; the χ and ω areas are both inflated. The inferior groove is wide and shallow, joining anteriorly to the hepatic groove. The carapace is densely ornamented by tubercles interspersed with irregular pits.
Pleon.—The pleonal segments are represented on specimen MCYCELHT 2/226 (Fig. 11.1); the tergites are subrectangular; the pleurae are subtriangular, directed backward, and bear a lateral bulge basally; s1 is short; s2 and s3 are of equal length; s4–6 are poorly preserved. The pleurae are ornamented by dense tubercles.
Thoracic appendages.—The P1 is partially preserved. The carpus of P1 is subcylindrical and laterally compressed, ornamented by dense tubercles. The propodus and dactylus are not preserved.
Material
PIN 5477/3784 and MCYCELHT 2/226 from the upper Callovian of Mikhailovtsement; PIN 5477/4537 from the upper Callovian of Gorenka.
Remarks
The specimens MCYCELHT 2/226 (Fig. 11.1), PIN 5477/3784 (Fig. 11.3, 11.4), and PIN 5477/4537 (Fig. 11.2) share characteristics of Erymidae as follows: the cervical, postcervical, hepatic, and inferior grooves are represented; short and shallow gastro-orbital groove beginning from the medial inflexion of the cervical groove; the postorbital area is delimited (not preserved on specimen PIN 5477/4537); the intercalated area is represented (specimen PIN 5477/3784, Fig. 11.3). (For a diagnosis of the family, see Devillez et al., Reference Devillez, Charbonnier and Pezy2018).
The studied material can be attributed to Eryma von Meyer, 1840 (von Meyer, Reference von Meyer1840a) because the postcervical groove ventrally joins to the branchiocardiac groove; the branchiocardiac groove ventrally joins to the hepatic groove; the hepatic groove is concavo-convex, forming the χ and ω areas; the orbital spine of the orbital row is represented. (For a diagnosis of Eryma, see Devillez et al., Reference Devillez, Charbonnier and Pezy2018; Devillez and Charbonnier, Reference Devillez and Charbonnier2021).
The studied specimens demonstrate peculiarities of E. quadriverrucatum as follows: the cervical groove is wide and strongly inclined; the gastro-orbital groove is short and deep; the postcervical and branchiocardiac grooves are deeply inclined; both χ and ω areas are inflated; the inferior groove is deep and wide; the ornamentation is represented by dense small tubercles surrounded with irregular depressions; the morphology of the pleonal somites is typical for the species (Devillez and Charbonnier, Reference Devillez and Charbonnier2021). It is important to note that the junction of the postcervical and branchiocardiac grooves of the studied specimens is situated at the mid-height of the carapace (Fig. 11.2–11.4). However, E. quadriverrucatum sensu Devillez and Charbonnier, Reference Devillez and Charbonnier2021 is characterized by a junction of the postcervical and branchiocardiac grooves situated very low on the carapace. Unfortunately, Devillez and Charbonnier did not study the type series but only original pictures of Trautschold (Devillez and Charbonnier, Reference Devillez and Charbonnier2021, fig. 11A, C). Thus, this feature of E. quadriverrucatum could be a misinterpretation of Trautschold's original work (Trautschold, Reference Trautschold1866) as the specimens depicted by Gerasimov were damaged but had crossing of the postcervical and branchiocardial grooves at the mid-height of the carapace (Gerasimov, Reference Gerasimov1955, pl. 8, figs. 1, 3; Devillez and Charbonnier, Reference Devillez and Charbonnier2021, fig. 11C, D). According to Devillez and Charbonnier (Reference Devillez and Charbonnier2021), the type age of E. quadriverrucatum is Tithonian; however, the age of the specimens could have been misinterpreted by Trautschold. Thus, we propose here the emended diagnosis for E. quadriverrucatum from the upper Callovian of Ryazan region.
The studied material differs from other Eryma species from the Middle and Late Jurassic by the following features: both χ and ω areas are inflated while only one or neither is inflated for E. compressum (Eudes-Deslongchamps, Reference Eudes-Deslongchamps1842), E. georgeii Carter, Reference Carter1886, E. jungostrix Feldmann and Titus, Reference Feldmann and Titus2006, E. lerasi (Étallon, Reference Étallon1861), E. mandelslohi (von Meyer, Reference von Meyer1840b), and E. ventrosum (von Meyer, Reference von Meyer1835); the carapace is ornamented by tubercles with irregular depressions while there are depressions only for E. mandelslohi and E. punctatum Oppel, Reference Oppel1861, tubercles with crescent-shaped depressions for E. compressum (Eudes-Deslongchamps, Reference Eudes-Deslongchamps1842), E. lerasi, and E. ventrosum, and tubercles only for E. jungostrix, E. osciensis Garassino et al., Reference Garassino, Bahrami, Yazdi and Vega2014, and E. westphali Schweigert et al., Reference Schweigert, Dietl and Röper2000. E. quadriverrucatum differs from E. ornatum (Quenstedt, Reference Quenstedt1858) and E. punctatum in that the postcervical and branchiocardiac grooves are crossing at the level of the gastro-orbital groove (crossing lower for E. ornatum and E. punctatum), and the postcervical groove has a ventral extension (absent for E. ornatum and E. punctatum). E. quadriverrucatum also differs from E. westphali by the spineless rostrum and from E. modestiforme (von Schlotheim, Reference von Schlotheim1822) by the rougher ornamentation of the carapace (despite the fine ornamentation for E. modestiforme).
Eryma sp.
Figure 12.1–12.6
Description
The specimens PIN 5477/4539 (Fig. 12.4, 12.5) and PIN 5477/4953 (Fig. 12.1–12.3) expose the isolated P1 chelae. The propodus is slightly laterally compressed (propodus length without index is ~12 mm; width is ~8 mm); the dorsal and ventral margins are narrow and carinate; the outer surface is slightly globose. The dactylar bulge is narrow and delimited by a deep groove. The fingers are elongate and equal in length; the index is strongly incurved at its basal part; the dactylus is almost straight and triangular in cross section. The ornamentation is represented by tubercles preceded with crescent-shaped depressions.

Figure 12. (1–6) Eryma sp.: (1–3) P1 chela from Gorenka, upper Callovian (PIN 5477/4953, dusted with ammonium chloride): (1) right ventral view, (2) right dorsal view, (3) basal view; (4, 5) P1 chela from Gorenka, upper Callovian (PIN 5477/4539, dusted with ammonium chloride): (4) left ventral view, (5) left dorsal view; (6) P1 chela from Gorenka, upper Callovian (PIN 5477/4538, dusted with ammonium chloride), left dorsal view. Stenodactylina sp.: (7) P1 finger from Gorenka, upper Callovian (PIN 5477/4541, dusted with ammonium chloride), lateral view. (8, 9) Stenodactylina insignis (Oppel, Reference Oppel1862), P1 chela from Mikhailovtsement, upper Callovian (PIN 5477/4540): (8) left dorsal view, (9) left ventral view. dac = dactylus of P1 chela; db = dactylar bulge; in = index of P1 chela; pr = propodus of P1. Scale bars = 10 mm.
The specimen PIN 5477/4538 (Fig. 12.6) exposes the isolated P1 chela as well as other specimens. The propodus is subrectangular and laterally compressed, with narrow inner and outer margins. The dactylar bulge is narrow and delimited by a deep groove. The P1 fingers are thin, poorly preserved; the dactylus is subtriangular in cross section; the occlusal margin of the dactylus bears large closely spaced teeth. The ornamentation is represented by tubercles without depressions.
Material
PIN 5477/4538, PIN 5477/4539, and PIN 5477/4953 from the upper Callovian of Gorenka.
Remarks
The studied material can be attributed to family Erymidae and genus Eryma on the basis of the subrectangular propodus laterally compressed, bearing a narrow dactylar bulge; thin fingers; uniform ornamentation (Devillez and Charbonnier, Reference Devillez and Charbonnier2021). However, these features are not sufficient to identify the specimens on the species level with confidence.
The general shape of specimens PIN 5477/4539 (Fig. 12.4, 12.5) and 5477/4953 (Fig. 12.1–12.3) is similar to those of E. mandelslohi (Devillez and Charbonnier, Reference Devillez and Charbonnier2019, fig. 7E, F) and E. lerasi (Devillez and Charbonnier, Reference Devillez and Charbonnier2021, fig. 6B). However, E. mandelslohi is considered to possess a bulge near the index base, which is not observed on the discussed specimens (Devillez and Charbonnier, Reference Devillez and Charbonnier2019). In the case of E. lerasi, the chelae attributed to this species by Devillez and Charbonnier (Reference Devillez and Charbonnier2021) strongly vary. The specimens depicted in their fig. 6F–I well correspond to the species diagnosis, whereas the specimens in their fig. 6A, B have a shorter propodus, which is less compressed in fig. 6B and more rounded in fig. 6A compared with the other depicted chelae (Devillez and Charbonnier, Reference Devillez and Charbonnier2021). The holotype of E. gracilimanum Lahusen, Reference Lahusen1894 (Devillez and Charbonnier, Reference Devillez and Charbonnier2021, fig. 6B) is considered to originate from the Tithonian, which could be the author's misinterpretation (Lahusen, Reference Lahusen1894).
Genus Stenodactylina Beurlen, Reference Beurlen1928
Type species
Stenodactylina liasina Beurlen, Reference Beurlen1928 by original designation from Toarcian, Holzmaden, Baden-Württemberg, Germany.
Stenodactylina insignis (Oppel, Reference Oppel1862)
Figure 12.8, 12.9
Type material
Cast of the holotype MNHN.F.A24613, collection of Muséum national d'Histoire naturelle, Paris, France (Devillez and Charbonnier, Reference Devillez and Charbonnier2021, fig. 16A)
Description
The specimen PIN 5477/4540 exposes the isolated P1 chela. The propodus is subrectangular and strongly elongate (length ~30 mm; width ~15 mm), globose in cross section, with outer margin more convex than inner one; the dactylar bulge is wide, delimited by a groove; the fingers are thin, very long, equal in width and length (preserved part ~30 mm); the index is wide basally, narrow at its middle and distal parts, having a strong inflexion at its basal third and sinuous distally; the dactylus is slightly incurved at its middle part; the occlusal margin has short conical teeth closely situated; the ornamentation of propodus is represented by small tubercles.
Material
PIN 5477/4540 from the upper Callovian of Mikhailovtsement.
Remarks
The specimen PIN 5477/4540 exposes the isolated chela (Fig. 12.8, 12.9) and is attributed to family Erymidae and genus Stenodactylina because the propodus is globose in cross section, the dactylar bulge is wide, the finger is slender, and the index is wide at its base and immediately narrowing distally.
Stenodactylina species are erected mostly on isolated chelae or carapaces (Feldmann et al., Reference Feldmann, Schweitzer and Haggart2020). The few species are known from the Callovian to the Oxfordian (Feldmann et al., Reference Feldmann, Schweitzer and Haggart2020; Devillez and Charbonnier, Reference Devillez and Charbonnier2019, Reference Devillez and Charbonnier2021): S. walkerae (Feldmann and Haggart, Reference Feldmann and Haggart2007) (Bajocian-Callovian, Canada), S. burgundiaca (Crônier and Courville, Reference Crônier and Courville2004) (Callovian, France), S. insignis (Oppel, Reference Oppel1862) (Oxfordian, France), and S. shotoverigiganti Devillez and Charbonnier, Reference Devillez and Charbonnier2021 (Oxfordian, UK). The specimen PIN 5477/4540 (Fig. 12.8, 12.9) is similar to S. insignis in the following: subrectangular propodus; index wide basally, with strong medial inflexion; short conical teeth on the occlusal margin; the ornamentation represented by small tubercles (Devillez and Charbonnier, Reference Devillez and Charbonnier2021, fig. 16A, C).
The specimen PIN 5477/4540 differs from S. burgundiaca in that the ornamentation is represented by small tubercles only (tubercles with crescent-shaped depressions for S. burgundiaca). Comparisons with S. walkerae and S. shotoverigiganti are quite difficult as those species are known only by isolated carapaces (Devillez and Charbonnier, Reference Devillez and Charbonnier2019).
Stenodactylina sp.
Figure 12.7
Description
The specimen PIN 5477/4541 exposes the isolated finger of P1 chela. The finger is large (preserved part is ~20 mm long), slender, slightly inflected basally, rounded in cross section, ornamented by small tubercles; the occlusal margin bears short, сlose, conical teeth.
Material
PIN 5477/4541 from the upper Callovian of Gorenka.
Remarks
We attribute the specimen PIN 5477/4541 to Stenodactylina because of its size, form, and ornamentation, which well correspond to those of the specimen PIN 5477/4540 fingers, whereas all the Eryma remains from the studied deposits are far smaller. Due to the lack of diagnostic features, we propose the combination Stenodactylina sp. for the specimen PIN 5477/4541.
Discussion
Although only three species of decapods, Archeosolenocera sp., Eryma quadriverrucatum Trautschold, Reference Trautschold1866, and Mecochirus sp., have been reported from the Callovian to the Oxfordian of European Russia, our study revealed a diverse assemblage of the crustaceans in the Ryazan region, which includes at least seven species from the four families (Solenoceridae, Mecochiridae, Glypheidae, and Erymidae). Among them, five taxa—Eumorphia sp., Glypheopsis aff. G. etalloni (Oppel, Reference Oppel1861), Eryma aff. E. ventrosum (von Meyer, Reference von Meyer1835), E. ornatum (Quenstedt, Reference Quenstedt1858), and Stenodactylina insignis (Oppel, Reference Oppel1862)—have never been previously acknowledged within the locations in European Russia.
Erymids of the Russian Middle Jurassic were traditionally attributed to E. quadriverrucatum Trautschold, Reference Trautschold1866 (Gerasimov, Reference Gerasimov1955; Gerasimov et al., Reference Gerasimov, Mitta, Kochanova and Tesakova1996), the presence of which in the studied deposits is here confirmed. The two other Eryma species, E. aff. E. ventrosum (von Meyer, Reference von Meyer1835) and E. ornatum (Quenstedt, Reference Quenstedt1858), are much less abundant, and the latter is a single decapod species observed in the Koenigii zone of the lower Callovian. We also report about an occurrence of Stenodactylina insignis (Oppel, Reference Oppel1862) in the deposits of Mikhailovtsement and Gorenka quarries. S. insignis is usual in the Callovian of France, but it has never been reported from European Russia. The new records of E. ornatum and S. insignis extend their area to the east and outline a faunistic exchange between decapod fauna of European Russia and those of Western Europe in the Callovian and the lower Oxfordian.
Mecochiridae are the second-most abundant family after Erymidae, previously noted in the Saratov region (Tesakova, Reference Tesakova2008). However, in the Moscow depression, they were overlooked by collectors and scientists for a long time, probably due to their small size. Most mecochirids are considered to be either epibenthic or half-burrowing animals (Simpson and Middleton, Reference Simpson and Middleton1985; Robin et al., Reference Robin, Charbonnier, Merle, Simpson, Petit and Fernandez2016), although in some works they are considered to be endobenthic (Förster, Reference Förster1971; Etter, Reference Etter2004). In Mikhailovtsement, Eumorphia sp. fossils are the most numerous in thin clay layers with phosphatized oolites. The same preservation is typical for Cretaceous Atherfieldastacus and Meyeria (Simpson and Middleton, Reference Simpson and Middleton1985; Robin et al., Reference Robin, Charbonnier, Merle, Simpson, Petit and Fernandez2016), which were associated with muddy substrates of the sublittoral zone. This condition is also specific for several localities in France containing Callovian crustacean remains (Crônier and Courville, Reference Crônier and Courville2004).
In contrast with Erymidae and Mecochiridae, glypheids are rare in the Jurassic of Moscow depression. Here we report first occurrences of glypheids in the middle and upper Callovian of European Russia. The lifestyle of extinct glypheids is considered to be similar to that of their living relatives (Boisselier-Dubayle et al., Reference Boisselier-Dubayle, Bonillo, Cruaud, Couloux, de Forges and Vidal2010; Charbonnier et al., Reference Charbonnier, Pérès and Letenneur2012, Reference Charbonnier, Audo, Barriel, Garassino, Schweigert and Simpson2014a), Neoglyphea inopinata Forest and de Saint Laurent, Reference Forest and de Saint Laurent1975 and Laurentaeglyphea neocaledonica (Richer de Forges, Reference Richer de Forges2006). Modern glypheids are either epi- or endobenthic and occur mostly on the muddy substrates or in burrows at depths of 180–260 and 360–530 m, respectively (Forest, Reference Forest2006; Charbonnier et al., Reference Charbonnier, Pérès and Letenneur2012).
The single occurrence of solenocerid Archeosolenocera sp., previously reported by Shmakov (Reference Shmakov2016) as Archeosolenocera straeleni Carriol and Riou, Reference Carriol and Riou1991 in the Mikhailovtsement quarry, Ryazan region, is quite surprising. Solenoceridae have become quite an abundant group recently, yet they were represented, until now, by three samples of extant genera (Alencar et al., Reference Alencar, Pinheiro, Saraiva, Oliveira and Santana2018) in the fossil record. Among them, only Pryorhyncha feitosai Alencar et al., Reference Alencar, Pinheiro, Saraiva, Oliveira and Santana2018 from the Cretaceous of Brazil was found in shallow-water deposits (Alenсar et al., Reference Alencar, Pinheiro, Saraiva, Oliveira and Santana2018). The other two species, Archeosolenocera straeleni and Eogordonella iranianiensis Garassino et al., Reference Garassino, Bahrami, Yazdi and Vega2014, are considered to inhabit the soft surfaces in deep water (Charbonnier et al., Reference Charbonnier, Vannier, Hantzpergue and Gaillard2009; Garassino et al., Reference Garassino, Bahrami, Yazdi and Vega2014; Jauvion et al., Reference Jauvion, Charbonnier and Bernard2017). Moreover, A. straeleni is endemic to La Voulte, the deposits of which originate from the hydrothermal vent (Charbonnier et al., Reference Charbonnier, Vannier, Hantzpergue and Gaillard2009; Reference Charbonnier, Audo, Caze and Biot2014b). Such specific habitat does not correspond to typical conditions in the Middle Russian Sea (Sahagian et al., Reference Sahagian, Pinous, Olferiev and Zakharov1996; Tesakova et al., Reference Tesakova, Shurupova and Ustinova2017) and might reflect the lower specificity of the genus Archeosolenocera to a particular depth. Among recent decapods, eurybathic distribution of a particular genus or even species is not exceptional (Fortibuoni et al., Reference Fortibuoni, Bahri, Camilleri, Garofalo, Gristina and Fiorentino2010; Linse et al., Reference Linse, Bohrmann and Sigwart2019; Rodriguez-Flores et al., Reference Rodríguez-Flores, Macpherson, Schnabel, Ahyong, Corbari and Machordom2022).
Whereas occurrences of crustaceans in the lower and middle Callovian of the region are still occasional, the upper Callovian and lower Oxfordian deposits contain abundant and relatively rich decapod assemblage (Table 2). An increase in the number of crustacean remains is more probably connected with good preservation conditions during this time rather than with real diversity changes. The modern stratigraphic data (Sahagian et al., Reference Sahagian, Pinous, Olferiev and Zakharov1996; Tesakova and Shurupova, Reference Tesakova and Shurupova2018) indicate the average depth of the sea in European Russia was 40 m or less by the end of the Callovian. The oolites with the crustacean remains were formed during a regression phase by slow sedimentation (Stepanov et al., Reference Stepanov, Kazansky, Kiselev, Kosareva, Rogov, Tesakova, Shchepetova and Shurupova2019) and were the most abundant in the upper sublittoral zone (Sahagian et al., Reference Sahagian, Pinous, Olferiev and Zakharov1996). Therefore, the Callovian decapod assemblage of the Mikhailovtsement and Gorenka quarries (Table 2) was composed mostly of epibenthic species and existed in the sublittoral zone on soft substrate. This assumption is confirmed by a specific ostracod composition (Tesakova, Reference Tesakova2008; Tesakova et al., Reference Tesakova, Shurupova and Ustinova2017; Tesakova and Shurupova, Reference Tesakova and Shurupova2018) and the presence of a rich assemblage of burrowing bivalves, such as Pholadomya Sowerby, Reference Sowerby1823, Gresslya Agassiz, Reference Agassiz1843, Pleuromya Agassiz, Reference Agassiz1842, and Trigonia Bruguière, Reference Bruguière1789, in the same localities. The decapod community of the Ryazan region is quite similar to the West European Callovian–Oxfordian faunas on the generic level. However, at least three observed species are specific for the region. The most abundant decapod observed only in European Russia, Eryma quadriverrucatum, is probably endemic for the area. Eryma aff. E. ventrosum and Glypheopsis aff. G. etalloni demonstrate significant differences from the European populations of those species. However, their statuses, together with those of Archeosolenocera sp. and Eumorphia sp., need further clarification. The relatively poor taxonomical composition of studied localities compared with the European Jurassic Lagerstätten (Van Straelen, Reference Van Straelen1925; Etter, Reference Etter, Bottjer, Etter, Hagadorn and Tang2002; Garassino and Schweigert, Reference Garassino and Schweigert2006; Charbonnier et al., Reference Charbonnier, Vannier, Hantzpergue and Gaillard2009; Odin et al., Reference Odin, Charbonnier, Devillez and Schweigert2019) might be due to the limited number of studies and the problems with preservation of large crustaceans. Most of the remains are extremely damaged or fragmented and preserved only in the small nodules; thus, the specific geological conditions created a bias to the lesser forms, lacking such groups as Aegeridae Burkenroad, Reference Burkenroad1963, Coleiidae van Straelen, Reference Van Straelen1925, and Polychelidae Wood-Mason, Reference Wood-Mason1874. The biodiversity of decapods in the studied area is still incomparable with those of Solnhofen (~70 crustacean species) and La Voulte (60 species) Lagerstätten (Garassino and Schweigert, Reference Garassino and Schweigert2006; Charbonnier et al., Reference Charbonnier, Vannier, Hantzpergue and Gaillard2009; Jauvion et al., Reference Jauvion, Charbonnier and Bernard2017). However, the occurrence of some other groups there, e.g., Prosopidae von Meyer, Reference von Meyer1860 and Tanidromitidae crabs (Crônier and Boursicot, Reference Crônier and Boursicot2009) mentioned by Gerasimov, Reference Gerasimov1955, is expected. Future efforts will shed more light on the decapod diversity of the Middle Russian Sea during the Callovian.
Table 2. List of the revealed taxonomic assemblages for the studied localities.

Conclusions
Seven species of Decapoda belonging to four families are observed in the localities of the Ryazan region, Central European Russia. Among them, four taxa (Glypheopsis aff. G. etalloni, Eryma ornatum, E. aff. E. ventrosum, and Stenodactylina insignis) are newly recorded in the Jurassic of the Ryazan region. For E. quadriverrucatum, the emended diagnosis is offered. In addition, the morphology and status of Archeosolenocera sp., recorded as A. straeleni by Shmakov (Reference Shmakov2016), and Eumorphia sp., recorded as Mecochirus sp. by Tesakova (Reference Tesakova2008), are discussed. The crustacean assemblage is composed mostly of small decapods that inhabited the shallow sublittoral zone with a soft substrate. The late Callovian–early Oxfordian faunistic complex of Gorenka and Mikhailovtsement includes several specific taxa absent in Western Europe. The hidden decapod diversity reflects a poor knowledge about the regional fauna, which probably includes some more groups overlooked by researchers.
Acknowledgments
We are very grateful to S.V. Bagirov and G.V. Zakharenko (Paleontological Institute, Moscow, Russia) for providing equipment and advice on photography and image processing. We cordially thank R. Feldmann (Kent State University, USA), J. Devillez, and S. Charbonnier (both Muséum national d'Histoire naturelle, Paris, France) for consultations on crustacean systematics and primary identification of the material. We declare our gratitude to M.A. Rogov, who kindly supplied stratigraphic data on Nikitino and gave advice on the geological structure of the other localities. We also appreciate I.A. Gaintsev, T.V. Gaintseva, A.Yu. Ganshkevich, L.V. Kulagina, S.B. Kulikova, K.O. Nazarov, and other participants in the Moscow Paleontological Society who collected the decapod remains. We express a separate gratefulness to D.B. Kucher (MCYCELHT) for access to their collections and L.V. Kulagina for immeasurable assistance with graphic content.
Declaration of competing interests
The authors declare no competing interests.














