



Introduction
Behavioral regulation is a complex socio-emotional executive function that involves inhibitory control, cognitive flexibility, and emotion control processes. Inhibitory control is the ability to suppress interfering distractions and prepotent motor responses (Diamond, Reference Diamond2013; Nigg, Reference Nigg2000). Cognitive flexibility, which is often measured using set-shifting, is the readiness with which one can switch from one task or mindset to another (Armbruster et al., Reference Armbruster, Ueltzhöffer, Basten and Fiebach2012; Diamond, Reference Diamond2013). Finally, emotion control is the process by which we influence the emotions we experience, when we experience them, and how we experience and express them (Gross, Reference Gross2002; Ochsner et al., Reference Ochsner, Silvers and Buhle2012).
Many children with neurodevelopmental conditions, including attention-deficit/hyperactivity disorder (ADHD) and developmental coordination disorder (DCD), have trouble regulating their behavior (Green & Payne, Reference Green and Payne2018; Posner et al., Reference Posner, Kass and Hulvershorn2014; Shaw et al., Reference Shaw, Stringaris, Nigg and Leibenluft2014; Tal Saban et al., Reference Tal Saban, Ornoy and Parush2014). They may be sensitive to external affective cues, making it hard for them to ignore distractions and follow instructions given by teachers or parents (Blair & Raver, Reference Blair and Raver2015; Diamond, Reference Diamond2013; Rosen et al., Reference Rosen, Walerius, Fogleman and Factor2015). They may also display frequent and intense shifts in emotions, and have trouble recovering from negative events (Rosen et al., Reference Rosen, Walerius, Fogleman and Factor2015). This struggle with behavioral regulation not only impacts children’s social relationships and performance at school, but also results in greater daily-life and mental health challenges overall (Barkley & Fischer, Reference Barkley and Fischer2010; Spencer et al., Reference Spencer, Faraone, Surman, Petty, Clarke, Batchelder and Biederman2011).
In children and young adults with ADHD (Barkley, Reference Barkley1997; Fischer et al., Reference Fischer, Barkley, Smallish and Fletcher2005), up to 50% have difficulty regulating their behavior and display high levels of emotional lability (Becker et al., Reference Becker, Steinhausen, Baldursson, Dalsgaard, Lorenzo and Ralston2006; Sobanski et al., Reference Sobanski, Banaschewski, Asherson, Buitelaar, Chen and Franke2010; Stringaris & Goodman, Reference Stringaris and Goodman2009). Evidence of treatment success with medication is limited (Lenzi et al., Reference Lenzi, Cortese, Harris and Masi2018), and many clinical trials have failed to address the difficulties in behavioral regulation that have been associated with ADHD in children (Posner et al., Reference Posner, Kass and Hulvershorn2014; Shaw et al., Reference Shaw, Stringaris, Nigg and Leibenluft2014). A handful of studies also suggest that children with DCD, a neurodevelopmental condition that is characterized by impaired motor coordination that significantly interferes with activities of daily living, school performance, as well as leisure and play activities (American Psychiatric Association, 2013), may have problems with behavioral regulation (Crane et al., Reference Crane, Sumner and Hill2017; Rahimi-Golkhandan et al., Reference Rahimi-Golkhandan, Steenbergen, Piek and Wilson2014; Rodriguez et al., Reference Rodriguez, Wade, Veldhuizen, Missiuna, Timmons and Cairney2019; van den Heuvel et al., Reference van den Heuvel, Jansen, Reijneveld, Flapper and Smits-Engelsman2016). To date, research that has examined behavioral regulation in pediatric populations has focused on “pure” neurodevelopmental conditions, including ADHD or DCD, and has not rigorously screened participants for comorbidities, although they frequently occur (Dewey et al., Reference Dewey, Kaplan, Crawford and Wilson2002; Fliers et al., Reference Fliers, Franke, Lambregts-Rommelse, Altink, Buschgens, Nijhuis-van der Sanden and Buitelaar2009). As such, closer examination of a neurodiverse group of children with ADHD, DCD, ADHD-DCD, and typically developing (TD) children will provide us with a better understanding of the spectrum of behavioral regulation.
Reliable brain-based markers of ADHD or DCD that support diagnostic phenotypes have been elusive due to the heterogeneity of these conditions. Examining the spectrum of expression of a specific feature, such as behavioral regulation, transdiagnostically may be more promising in identifying brain-based markers of these conditions (Ameis et al., Reference Ameis, Lerch, Taylor, Lee, Viviano, Pipitone and Anagnostou2016; Lake et al., Reference Lake, Finn, Noble, Vanderwal, Shen, Rosenberg and Constable2019; Uddin et al., Reference Uddin, Dajani, Voorhies, Bednarz and Kana2017). The neural substrates of behavioral regulation have been extensively studied in neurotypical adults (Morawetz et al., Reference Morawetz, Bode, Derntl and Heekeren2017) and adults with affective disorders (Picó-Pérez et al., Reference Picó-Pérez, Radua, Steward, Menchón and Soriano-Mas2017), but less is known about the neural expression of behavioral regulation in pediatric populations. A handful of studies with relatively small sample sizes (N < 50) in children with ADHD have shown that behavioral regulation is associated with alterations in the prefrontal cortex (PFC), including orbitofrontal cortex (OFC), and the anterior cingulate cortex (ACC), as well as in limbic and reward areas such as the amygdala, insula, and accumbens (Hulvershorn et al., Reference Hulvershorn, Mennes, Castellanos, Di Martino, Milham, Hummer and Roy2014; Passarotti et al., Reference Passarotti, Sweeney and Pavuluri2010; Posner et al., Reference Posner, Maia, Fair, Peterson, Sonuga-Barke and Nagel2011, Reference Posner, Rauh, Gruber, Gat, Wang and Peterson2013). Considering that problems in behavioral regulation are common in children with ADHD (Posner et al., Reference Posner, Kass and Hulvershorn2014; Shaw et al., Reference Shaw, Stringaris, Nigg and Leibenluft2014) and also reported in children with DCD (Crane et al., Reference Crane, Sumner and Hill2017; Rahimi-Golkhandan et al., Reference Rahimi-Golkhandan, Steenbergen, Piek and Wilson2014; Rodriguez et al., Reference Rodriguez, Wade, Veldhuizen, Missiuna, Timmons and Cairney2019; van den Heuvel et al., Reference van den Heuvel, Jansen, Reijneveld, Flapper and Smits-Engelsman2016), and the widespread repercussions suboptimal behavioral regulation can have throughout childhood and into adulthood, systematic characterization of the interactions of these areas with the rest of the brain, or their functional connectivity (FC), transdiagnostically, has enormous potential for the diagnosis and development of individually tailored treatment for behavioral regulation difficulties in children with various neurodevelopmental conditions (Shaw et al., Reference Shaw, Stringaris, Nigg and Leibenluft2014). Examining distributed FC patterns, that is, FC patterns spanning across multiple brain networks, provides a more holistic perspective of the associations between brain functions and behaviors than can be gleaned from analyzing brain activity or FC of a single region alone. Indeed, looking at FC patterns transdiagnostically and across multiple brain networks has been useful in improving our understanding of inattention and hyperactivity in children with and without ADHD (Elton et al., Reference Elton, Alcauter and Gao2014; Rosenberg et al., Reference Rosenberg, Finn, Scheinost, Papademetris, Shen, Constable and Chun2016), and behavioral regulation in children with and without Autism Spectrum Disorder (Rohr et al., Reference Rohr, Kamal and Bray2020).
The primary aim of this study was to provide a comprehensive picture of the FC signatures associated with behavioral regulation. To accomplish this, we investigated the FC signatures underlying behavioral regulation transdiagnostically in a unique cohort of TD children, children with ADHD, children with comorbid ADHD-DCD and children with DCD without any known comorbidities. We used the behavioral regulation index score on the Behavior Rating Inventory of Executive Function (BRIEF), a parent report measure, as our primary outcome and examined associations between behavioral regulation and 10 prefrontal, limbic and striatal regions of interest in resting-state fMRI data of TD children and children with ADHD, DCD, or ADHD-DCD. Behaviorally, we hypothesized that children with a neurodevelopmental condition would evidence more problems in behavioral regulation than TD children. Neurally, we hypothesized that the FC of prefrontal, limbic, and striatal regions would show transdiagnostic associations with behavioral regulation.
Methods and materials
This research was conducted in accordance with the Declaration of Helsinki for experiments involving human subjects. It was approved by the Conjoint Health Research Ethics Board of the University of Calgary. Written consent and verbal assent were obtained from parents or guardians, and participants, respectively.
Participants
Recruitment and screening
Participants were recruited from local schools and through community advertisements in locations such as hospitals and physician’s offices in Calgary, Alberta, Canada. TD children and children diagnosed with ADHD, DCD, or ADHD-DCD, as well as children with attention and/or motor difficulties, were eligible, provided they had not been diagnosed with another neurodevelopmental or psychiatric disorder, a neurological, metabolic or genetic condition, and were not born preterm (<36 weeks) or with very low birth weight (<1500 g). Potential participants were screened for contraindications for MRI and other medical problems that would prevent participation.
Neuropsychological assessment for diagnosis
Recruited participants who met the above criteria were invited to participate in a detailed neuropsychological assessment. Data were collected over several years. Children were classified as ADHD or DCD in keeping with the Diagnostic and Statistical Manual of Mental Disorders - Fourth Edition Text Revision (DSM-IV-TR) (American Psychiatric Association, 2000), because the DSM-IV was still the standard diagnostic manual in Canada when data collection began and these criteria were used throughout the study. Parents completed the ADHD module of the Diagnostic Interview for Children and Adolescents – IV (DICA-IV) computerized interview (Reich et al., Reference Reich, Weltner and Herjanic1997), which evaluates inattention and hyperactivity on several dimensions of behavior and activities of daily living. A score of “1” indicates significant impairment with respect to attention (A criterion) or hyperactivity (B criterion), and a score of “0” indicates that there is no evidence of symptoms. On the Conners’ Parent Rating Scale – Revised (CPRS-R; Conners et al., Reference Conners, Sitarenios, Parker and Epstein1998), parents rate a range of behaviors associated with ADHD and behavior problems in children. The mean T-score is 50 (SD = 10) and children with scores above 60 can be indicative of ADHD. Children were classified as ADHD if they met the diagnostic criteria on the DICA-IV (Reich et al., Reference Reich, Weltner and Herjanic1997), or had a T-score above the 95th percentile on the CPRS-R (Conners et al., Reference Conners, Sitarenios, Parker and Epstein1998) and were diagnosed by a physician as having ADHD based on DSM-IV-TR criteria. The Movement Assessment Battery for Children - Second Edition (MABC-II) is a valid standardized motor assessment that evaluates motor performance across three domains: manual dexterity, aiming and catching and balance skills (Schoemaker et al., Reference Schoemaker, Niemeijer, Flapper and Smits-Engelsman2012; Van Waelvelde et al., Reference Van Waelvelde, Peersman, Lenoir and Smits Engelsman2007). The mean standard score on this measure is 10 (SD = 3) and higher scores on this measure indicate better performance. The Developmental Coordination Questionnaire (Wilson et al., Reference Wilson, Kaplan, Crawford, Campbell and Dewey2000) is a valid parent report that can be used to screen for motor problems in children that affect daily functioning. Higher scores on this measure indicate better motor functioning. Children were classified as DCD if they displayed an impairment in motor function (i.e., scored ≤ 16th percentile on the MABC-II) (Henderson et al., Reference Henderson, Sudgen and Barnett2007), were reported by their parents as exhibiting motor difficulties that interfered significantly with daily functioning on the Developmental Coordination Questionnaire (Wilson et al., Reference Wilson, Kaplan, Crawford, Campbell and Dewey2000), did not evidence a visual impairment or other neurological/medical condition that would affect movement and did not display an intellectual impairment as evidenced by performance on a standardized measure of cognitive function, i.e. the Wechsler abbreviated scale of intelligence (WASI) (Wechsler, Reference Wechsler1999). The WASI (Wechsler, Reference Wechsler1999) is a short standardized assessment that provides a valid and reliable (reliability of 0.90) measure of intelligence. It has a mean of 100 (SD = 15) and higher scores indicate a better performance. Participants completed all four WASI subtests (Block Design, Vocabulary, Matrix Reasoning and Similarities). Handedness was determined based on the preferred hand identified and used by the child when performing fine motor tasks on standardized measures of motor function (i.e., MABC-II) (Henderson et al., Reference Henderson, Sudgen and Barnett2007). Children meeting criteria for both ADHD and DCD were classified as ADHD-DCD. Children in the TD group did not meet criteria for ADHD or DCD. Children who were prescribed stimulant treatment for ADHD were asked to refrain from taking their medication on the day they underwent MRI scanning.
Final sample
A total of 149 participants who met criteria underwent resting state fMRI. Of these, 6 did not complete the diagnostic assessment measures; 1 (ADHD-DCD) was found to have a diagnosis of Autism Spectrum Disorder; 14 (6 TD, 3 DCD, 3 ADHD, 2 ADHD-DCD) did not complete the cognitive assessment; and 4 (1 ADHD-DCD, 1 ADHD, 2 DCD) did not complete the MRI scan. Of the remaining participants, nine had excessive head motion on their fMRI scan (>5 mm maximum absolute displacement). Participants’ data were further evaluated for outliers on behavioral measures, defined as > 3 SD from the mean. No participant was excluded due to this criterion. The final sample consisted of 115 participants; characteristics are provided in Table 1.
Table 1. Participant characteristics

Note. Means and standard deviations (in brackets) are provided for the total sample, as well as for TD participants and participants with ADHD, DCD, and ADHD-DCD and the children without and with ADHD, separately. Motion (mm) refers to the absolute maximum displacement at any timepoint in the resting-state fMRI scan prior to motion mitigation and denoising procedures. N = number of participants; FSIQ = Full-Scale Intelligence Quotient; Behavioral regulation index scores, as well as scores on the subscales (inhibition, shifting, and emotion control) are given as T-scores. †denotes a significant difference between children with and without ADHD at p < .05.
Behavioral regulation assessment
Behavioral regulation was assessed with the BRIEF (Gioia et al., Reference Gioia, Isquith, Guy and Kenworthy2000), a standardized parent report measure of executive function behaviors for children aged 5–18 years. The BRIEF provides a composite behavioral regulation index score, which includes three subdomains of behavioral regulation: “inhibit”, “shift”, and “emotion control”. The “inhibit” subscale assesses the ability to resist impulses and to stop one’s own behavior” (sample item: “acts wilder or sillier than others in groups (birthday parties, recess)”). The “shift” subscale assesses the ability to move freely from one situation, activity, or problem to another; to tolerate change, and to switch or alternate attention (sample item: “resists or has trouble accepting a different way to solve a problem with schoolwork, friends, chores, etc.”). Finally, the “emotion control” subscale assesses the ability to regulate emotional responses appropriately (sample item: “overreacts to small problems”). Together, scores in these subscales make up the behavioral regulation index score. Normed T-scores with a mean of 50 (SD = 10) were used in the analyses, with higher scores indicating more problems in behavioral regulation.
MRI data acquisition parameters
Data were acquired at the Seaman Family MR Research Centre at the University of Calgary across two MRI systems due to a system upgrade. Sixty-seven scans were collected on a 3T GE Signa VH/i (Waukesha, WI) with an eight-channel phased-array radiofrequency head coil and 48 scans were collected on a GE 750 with an eight-channel phased-array head coil. Children were instructed to keep their eyes on a fixation cross at the center of the screen. Functional images were acquired using a gradient-echo EPI sequence in 40 axial slices (120 volumes, TR = 2500 ms, TE = 30 ms, FA = 70, matrix size 64 × 64, voxel size 3.44 × 3.44 × 3 mm3; duration: 5 min) in the first round of acquisition, and in 26 axial slices (140 volumes, TR = 2500 ms, TE = 30 ms, FA = 70, matrix size 64 × 64, voxel size 3.44 × 3.44 × 4 mm3; duration: 5.8 min) in the second round of acquisition. Anatomical scans were acquired using a T1-weighted MPRAGE sequence (TR = 1000 ms, TE = 2.5 ms, FA = 18, voxel size 0.9 × 0.9 × 4 mm3 in the first round of acquisition and TR = 7.4 ms, TE = 3.1 ms, FA = 13, voxel size 1 × 1 × 0.8 mm3 in the second round of acquisition).
MRI data preprocessing
Data preprocessing used functions from FSL (Smith et al., Reference Smith, Jenkinson, Woolrich, Beckmann, Behrens, Johansen-Berg and Matthews2004) and AFNI (Cox, Reference Cox1996) and integrated “best-in-breed” tools for each preprocessing step covered in the workflow akin to the approach taken by fMRIprep (Esteban et al., Reference Esteban, Markiewicz, Blair, Moodie, Isik, Erramuzpe and Gorgolewski2019). The specific functions are denoted in brackets. Anatomical data were deobliqued (3drefit), oriented into FSL space (RPI) (3dresample) and skull-stripped (3dSkullStrip and 3dcalc). Functional data were also first deobliqued (3drefit) and oriented into FSL space (RPI) (3dresample). The pipeline further consisted of motion correction (MCFLIRT), skull-stripping (3dAutomask and 3dcalc), spatial smoothing (6 mm Gaussian kernel full-width at half-maximum) (fslmaths), grand-mean scaling (fslmaths), registration to the participant’s anatomical scan (FLIRT), and normalization to the McConnell Brain Imaging Center NIHPD asymmetrical (natural) pediatric template optimized for ages 4.5–18.5 years (Fonov et al., Reference Fonov, Evans, Botteron, Almli, McKinstry, Collins and Group2011) (FLIRT), followed by normalization to 2 × 2 × 2 mm MNI152 standard space (FLIRT).
Head motion and physiological confound mitigation procedure
A four-step process was used to address motion and physiological confounds in the data. First, motion estimates derived from the preprocessing were utilized to exclude participants with excessive head motion; scans were excluded if they exhibited >5 mm absolute maximum displacement. Second, AROMA was employed, an ICA-based cleaning method (Pruim et al., Reference Pruim, Mennes, van Rooij, Llera, Buitelaar and Beckmann2015), which allows for the retention of the remaining “true” neural signal within an affected volume (Kaufmann et al., Reference Kaufmann, Alnæs, Doan, Brandt, Andreassen and Westlye2017). AROMA is an automated procedure that uses a small but robust set of theoretically motivated temporal and spatial features (time series and power spectrum) to distinguish between “real” neural signals and motion artifacts. We chose a threshold that is conservative about what is retained (“aggressive”) to decrease the chance of false positives. Noise components identified by AROMA were removed from the data. Third, images were de-noised by regressing out the six motion parameters, as well as signal from white matter, cerebral spinal fluid and the global signal, as well as their first-order derivatives (Parkes et al., Reference Parkes, Fulcher, Yücel and Fornito2018). While there is currently no gold standard (Murphy & Fox, Reference Murphy and Fox2017) regarding the removal of the global signal, it was removed here based on evidence that it relates strongly to respiratory and other motion-induced signals, which persist through common denoising approaches including ICA and models that approximate respiratory variance (Power et al., Reference Power, Plitt, Gotts, Kundu, Voon, Bandettini and Martin2018). Motion (defined as each participant’s absolute maximum displacement) was substantially reduced following this procedure (before: 1.5 ± 1.2 mm; after: 0.07 ± 0.03 mm). As a final step, described below, head motion, defined as absolute maximum displacement, was included in the analysis models as a covariate of no interest. Including motions as a covariate in a regression model can reduce motion-related group differences (Power et al., Reference Power, Schlaggar and Petersen2015). This approach was chosen for our pediatric sample to minimize a residual influence of motion on the results as numerical differences in motion were noted among diagnostic groups.
Analysis of demographic, diagnostic, and behavioral measures
ANOVAs were used to examine differences in demographics and diagnostic measures among the four participant groups: TD children and children with ADHD, DCD, or ADHD-DCD. ANCOVAs were then used to assess differences in behavioral regulation, controlling for any observed differences in demographics among the four diagnostic groups as covariates of no interest. As described in the Results section below, there were no differences in behavioral regulation between TD children and children with DCD, and no differences between children with ADHD and ADHD-DCD; therefore, we focused the analysis on children with ADHD (ADHD and ADHD-DCD) versus children without ADHD (TD children and children with DCD). T-tests were utilized to assess differences in demographics, head motion and behavioral regulation between these groups. Finally, Pearson correlations were computed to assess the relationship between demographics, head motion and behavioral regulation scores. These analyses were carried out using SPSS 22 (Chicago, IL).
Analysis of fMRI data
To examine the associations between FC of the regions of interest and behavioral regulation scores across the brain for the entire sample of children, 10 regions were selected based on a well-known model of behavioral regulation (Ochsner et al., Reference Ochsner, Silvers and Buhle2012) and ADHD meta-analyses (Cortese et al., Reference Cortese, Castellanos, Eickhoff, D'Acunto, Masi, Fox and Eickhoff2016; Frodl & Skokauskas, Reference Frodl and Skokauskas2012; Hoogman et al., Reference Hoogman, Bralten, Hibar, Mennes, Zwiers, Schweren and Franke2017) (see Figure 1 and Table S1 for details). Each region’s FC map was then computed using AFNI. First, the average time course was extracted for each region (3dROIstats) and entered into a voxel-wise correlation with every other voxel in the brain using cross-correlation (3dfim+). Resultant whole-brain FC maps were normalized using Fisher’s r-to-z transform (z = .5[ln(1+r)-ln(1−r)]) for comparison across individuals (3dcalc). Group-level statistical testing was conducted with FLAME 1, a mixed-effects analysis in FSL’s FEAT using automatic outlier deweighing. In a regression analysis, the behavioral regulation index T-score was converted to a z-score and entered into a model that included z-scored age, FSIQ, sex, scanner and motion as nuisance covariates, to assess the association between FC and behavioral regulation across the entire sample of children. Voxel-wise thresholding was set at z-score >2.3, and cluster correction was conducted using Gaussian Random Field theory with p < .05. The p-values for these results were then Bonferroni-corrected for twenty comparisons (i.e., the number of seeds that were examined; significance set at p < .0025).

Figure 1. Seed regions of interest. To examine how FC associates with behavioral regulation scores across the brain and how FC differs between groups, 10 ROIs were selected in limbic areas (i.e., amygdala and insula), prefrontal areas (i.e., dorsolateral, dorsomedial, and ventromedial prefrontal cortex; orbitofrontal cortex and subgenual anterior cingulate cortex), and striatal areas (i.e., caudate, putamen, and accumbens). Regions were anatomically defined using probabilistic parcellation units provided through FSL with the Harvard-Oxford Atlas and thresholded at 50% probability, meaning any given voxel within the seed mask had a > 50% probability of lying within the specified region. Masks were binarized.
Assessment of specificity to behavioral regulation
To evaluate whether our correlation analyses captured behavioral regulation dimensionally or were driven by the categorical difference in scores due to ADHD diagnosis, we performed a post hoc correlation analysis accounting for diagnostic status through an added nuisance covariate.
Results
Sample characteristics
Characteristics for the sample are provided in Table 1 and results for all comparative tests on demographic, diagnostic and behavioral measures can be found in Tables S2–S4. There were no significant differences in behavioral regulation scores between TD children and children with DCD, or between children with ADHD and children with ADHD-DCD. Group comparisons were therefore carried out only on the combined groups of children with ADHD (ADHD and ADHD-DCD, n = 63) versus children without ADHD (TD and DCD, n = 52). Significant differences between children with and without ADHD existed in sex (p = .0002), IQ (p = .035) and the distribution across scanners (p = .006), but not in motion (neither before cleaning nor after; both p > .11). Adjusting for these covariates (i.e., sex, IQ, and distribution across scanners), results still showed significant differences between children with and without ADHD in behavioral regulation (p = .000054), reflecting greater challenges with behavioral regulation for children with ADHD. No correlations were observed between behavioral regulation and age, FSIQ or motion (neither before cleaning nor after; all p > .23).
Transdiagnostic functional connectivity associated with behavioral regulation
A total of eight FC patterns across four seeds were associated with behavioral regulation across all participants (Table 2) in the regression analysis. These were seeds in vmPFC, sgACC, OFC, and accumbens. FC associated with behavioral regulation followed four main patterns: (1) FC within medial-prefrontal areas; (2) FC between medial-prefrontal and lateral-prefrontal areas; (3) FC between medial-prefrontal areas and limbic-striatal areas; and (4) FC between accumbens and visual areas (Figure 2). Overall, greater behavioral regulation problems were associated with stronger negative FC, but also with weaker positive FC in 25% (n = 2) of the behavioral regulation-associated patterns.

Figure 2. Predominant FC patterns that associated with behavioral regulation scores across all participants. Stronger negative FC was associated with greater behavioral regulation problems between the right and left nucleus accumbens and bilateral visual cortex 1–2 (a; FC map and plot of left nucleus accumbens is shown), as well as between the right and left vmPFC and a cluster spanning left vlPFC and dlPFC (b; FC map and plot of right vmPFC is shown). Weaker positive FC was associated with greater behavioral regulation problems between left OFC and a cluster spanning right vACC and nucleus accumbens (c), as well as between left sgACC and a dorsal striatal cluster (d). Negative associations between FC and behavioral regulation scores are depicted in blue-light blue. Colored arrows with –/+ signs indicate the mean direction of FC between seed and cluster regions as identified via the seed’s FC map. Correlation plots show all values adjusted for age, IQ, sex, scanner, and motion. Results are corrected for multiple comparisons at p < .0025 (20 seeds, 10 in each hemisphere). ACC = anterior cingulate cortex; dlPFC = dorsolateral prefrontal cortex; R = right hemisphere; V1–V2 = visual cortex 1–2; vACC = ventral anterior cingulate cortex; vlPFC = ventrolateral prefrontal cortex; vmPFC = ventromedial prefrontal cortex.
Table 2. Associations between behavioral regulation index scores and FC across all participants
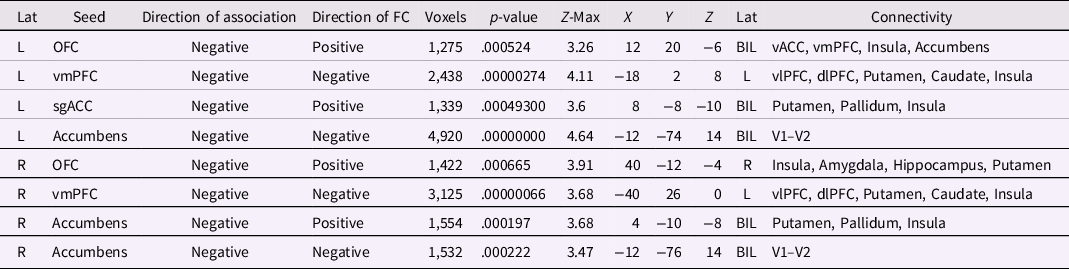
Note. BIL = bilateral; dlPFC = dorsolateral prefrontal cortex; dmPFC = dorsomedial prefrontal cortex; L = left; Lat = Laterality; OFC = orbitofrontal cortex; R = right; rACC = rostral anterior cingulate cortex; ROI = region of interest; sgACC = subgenual anterior cingulate cortex; V1–V2 = visual cortex 1–2; V2–V3 = visual cortex 2–3; vACC = ventral anterior cingulate cortex; vlPFC = ventrolateral prefrontal cortex; vmPFC = ventromedial prefrontal cortex; TP = temporal pole. Direction of Association refers to the direction of the association with behavioral regulation. Direction of FC refers to the direction of FC between seed region and connectivity cluster. Results are corrected for multiple comparisons at p < .0025 (20 seeds, 10 in each hemisphere).
Specificity to behavioral regulation
All FC patterns detected in the regression analysis remained associated with behavioral regulation after controlling for ADHD diagnosis, suggesting that these effects were not driven by diagnostic status.
Discussion
Poorer behavioral regulation is a known issue for children with neurodevelopmental conditions such as ADHD and is associated with greater daily-life challenges and an increased risk for psychiatric comorbidities (Barkley & Fischer, Reference Barkley and Fischer2010; Spencer et al., Reference Spencer, Faraone, Surman, Petty, Clarke, Batchelder and Biederman2011). In this study, which examined behavioral regulations across diagnostic groups (i.e., transdiagnostically), the strength of distributed patterns of FC among prefrontal, limbic, striatal, and visual brain areas was associated with children’s individual differences in behavioral regulation, and these associations remained significant after taking ADHD diagnostic status into account. Specifically, we found that FC within medial-prefrontal areas and FC between medial-prefrontal and limbic or striatal areas was significantly associated with behavioral regulation. Likewise, behavioral regulation was associated with FC between medial-prefrontal and lateral-prefrontal areas as well as with FC between reward and visual areas. However, children with a diagnosis of ADHD (i.e., children with ADHD or ADHD-DCD) had significantly more problems in behavioral regulation than TD children and children with “pure” DCD. These findings suggest that selected subsets of FC data involving frontostriatal, limbic, and visual pathways may have utility as brain-based signatures of behavioral regulation problems across children with and without ADHD despite significant differences in behavioral regulation scores.
Transdiagnostically, FC associated with behavioral regulation fell within four main seed regions − vmPFC, sgACC, OFC, and accumbens − and greater behavioral regulation problems tended to be associated either with weaker positive or with stronger negative FC. For instance, stronger negative FC between vmPFC and vlPFC/dlPFC associated with greater behavioral regulation problems, and this pattern existed bilaterally. vmPFC anatomically connects to dlPFC via vlPFC, and while individual differences in gray matter volume in vlPFC and dlPFC predicted regulatory success in a self-control study (Schmidt et al., Reference Schmidt, Tusche, Manoharan, Hutcherson, Hare and Plassmann2018), and functional activity in these regions was associated with an object’s attributed value (Hutcherson et al., Reference Hutcherson, Plassmann, Gross and Rangel2012), they may have distinct roles in behavioral regulation processes. For instance, the downregulation of cravings has been found to selectively modulate dlPFC activity, while the upregulation of cravings has been found to modulate vmPFC activity (Hutcherson et al., Reference Hutcherson, Plassmann, Gross and Rangel2012). vlPFC was functionally connected to vmPFC and dlPFC during both regulation processes, and it has been theorized that vlPFC may help to implement changes to the circuitry generated by the initiation of a behavioral regulation strategy (Hutcherson et al., Reference Hutcherson, Plassmann, Gross and Rangel2012). Refining the notion of distinct roles for the vmPFC and dlPFC further, it has been suggested that the vmPFC integrates affective valuations (made by amygdala and accumbens, rather than vmPFC itself) with inputs from prefrontal control centers like vlPFC and dlPFC that provide information about current behavioral goals (Hare et al., Reference Hare, Camerer and Rangel2009; Ochsner et al., Reference Ochsner, Silvers and Buhle2012). Thus, it seems reasonable to assume that this FC pathway between medial and lateral PFC may reflect a behavioral regulation process that integrates valuation and current behavioral goals.
Stronger negative FC between accumbens and primary visual areas also associated with greater behavioral regulation problems, and again the pattern existed bilaterally. FC between accumbens and primary visual areas has been observed during reward processing (Weiland et al., Reference Weiland, Welsh, Yau, Zucker, Zubieta and Heitzeg2013) and accumbens and visual areas have been jointly activated in reward-directed action and inhibition of action, (Le et al., Reference Le, Zhang, Zhornitsky, Wang and Li2020) and response to incentives (Gorka et al., Reference Gorka, Fuchs, Grillon and Ernst2018). Accumbens receives projections from dopamine-releasing neurons, making it rich in dopamine (Ikemoto, Reference Ikemoto2010). Dopamine is thought to code for learned associations and mediate approach behavior toward a reward; it is known to be actively involved in behavioral regulation tasks requiring cognitive flexibility, (Klanker et al., Reference Klanker, Feenstra and Denys2013) and plays an important role in processing rewarding and reinforcing stimuli (e.g., food) (Olsen, Reference Olsen2011) as well as in reward anticipation (Schuetze et al., Reference Schuetze, Rohr, Dewey, McCrimmon and Bray2017) and outcome prediction (Bray & O'Doherty, Reference Bray and O'Doherty2007; Schuetze et al., Reference Schuetze, Cho, Vinette, Rivard, Rohr, Ten Eycke and Bray2019). Reward and behavioral regulation are arguably linked, with the term cognitive reward control being used to describe the regulation of one’s behavior towards hedonic stimuli like food (Brandl et al., Reference Brandl, Le Houcq Corbi, Mulej Bratec and Sorg2019). This is especially true in children (Power et al., Reference Power, Olivera, Hill, Beck, Hopwood, Garcia and Hughes2016). Thus, it stands to reason that this may be a visual reward FC pathway used in responding to incentives, as well as in shifting of reward-directed action and inhibition of that action.
Greater behavioral regulation problems were also associated with weaker positive FC in two patterns associated with behavioral regulation. Both FC patterns centered on frontostriatal reward pathways repeatedly shown to be heavily affected in ADHD (Norman et al., Reference Norman, Carlisi, Christakou, Murphy, Chantiluke, Giampietro and Rubia2018). Both also involved the ACC and it should be noted that ACC FC is crucial in monitoring for potential conflicts and prepotent responses (Egner et al., Reference Egner, Etkin, Gale and Hirsch2008; Etkin et al., Reference Etkin, Prater, Hoeft, Menon and Schatzberg2010; Rohr et al., Reference Rohr, Villringer, Solms-Baruth, van der Meer, Margulies and Okon-Singer2016). Behavioral regulation has been associated with FC between the OFC and accumbens/vACC, and animal studies have shown that hemodynamic signals of, and neuronal projections between, OFC and accumbens are related to inhibition-related processes that are part of reinforcement learning (Groman et al., Reference Groman, Keistler, Keip, Hammarlund, DiLeone, Pittenger and Taylor2019; Werlen et al., Reference Werlen, Shin, Gastambide, Francois, Tricklebank, Marston and Walton2019). Behavioral regulation has also been associated with FC between sgACC and putamen/pallidum and activity in both structures has been found to be aberrant during reward prediction in obsessive-compulsive disorder (OCD) (Hauser et al., Reference Hauser, Iannaccone, Dolan, Ball, Hättenschwiler, Drechsler and Brem2017), a disorder often comorbid with ADHD and that like ADHD is a “disorder of control” (Brem et al., Reference Brem, Grünblatt, Drechsler, Riederer and Walitza2014). Further, volume in both structures has been found to be different in adult and pediatric individuals with ADHD (Frodl & Skokauskas, Reference Frodl and Skokauskas2012) and OCD (Ahmed et al., Reference Ahmed, Ras and Seedat2012; Gilbert et al., Reference Gilbert, Keshavan, Diwadkar, Nutche, Macmaster, Easter and Rosenberg2008).
Individual differences in behavioral regulation have been repeatedly found to be associated with individual features in FC (Ferri et al., Reference Ferri, Schmidt, Hajcak and Canli2016; Fitzgerald et al., Reference Fitzgerald, Klumpp, Langenecker and Phan2019; Rohr et al., Reference Rohr, Villringer, Solms-Baruth, van der Meer, Margulies and Okon-Singer2016). Taking individual differences into account can help expose the underlying neural substrates of complex cognitive skills, emotions and social competencies, and has proven useful in the investigation of both neurotypical (Goldfarb et al., Reference Goldfarb, Chun and Phelps2016; Rohr et al., Reference Rohr, Okon-Singer, Craddock, Villringer and Margulies2013, Reference Rohr, Dreyer, Aderka, Margulies, Frisch, Villringer and Okon-Singer2015; Vossel et al., Reference Vossel, Weidner, Moos and Fink2016) and clinical populations (Nebel et al., Reference Nebel, Eloyan, Nettles, Sweeney, Ament, Ward and Mostofsky2015; van Dongen et al., Reference van Dongen, von Rhein, O'Dwyer, Franke, Hartman, Heslenfeld and Buitelaar2015; von Rhein et al., Reference von Rhein, Cools, Zwiers, van der Schaaf, Franke, Luman and Buitelaar2015), as traits and abilities associated with neurodevelopmental conditions such as ADHD exist in the neurotypical population, falling on a spectrum (Matthews et al., Reference Matthews, Nigg and Fair2014; van Dongen et al., Reference van Dongen, von Rhein, O'Dwyer, Franke, Hartman, Heslenfeld and Buitelaar2015). Examination of individual differences also allows for more statistical power in studies that include children with neurodevelopmental conditions such ADHD and DCD, which often struggle with small, heterogenous samples (Fair et al., Reference Fair, Bathula, Nikolas and Nigg2012; Nigg, Reference Nigg2005; Sonuga-Barke et al., Reference Sonuga-Barke, Sergeant, Nigg and Willcutt2008; Uddin et al., Reference Uddin, Dajani, Voorhies, Bednarz and Kana2017).
Unlike several recent studies (Crane et al., Reference Crane, Sumner and Hill2017; Rahimi-Golkhandan et al., Reference Rahimi-Golkhandan, Steenbergen, Piek and Wilson2014; Rodriguez et al., Reference Rodriguez, Wade, Veldhuizen, Missiuna, Timmons and Cairney2019; van den Heuvel et al., Reference van den Heuvel, Jansen, Reijneveld, Flapper and Smits-Engelsman2016), we found no elevation of behavioral regulation scores in children with “pure” DCD. This may be because we rigorously screened for comorbid ADHD; up to 50% of children with DCD meet diagnostic criteria for ADHD but only 5% are diagnosed (McLeod et al., Reference McLeod, Langevin, Dewey and Goodyear2016). We also found that children with ADHD-DCD showed elevated scores on behavioral regulation; therefore, it is likely that the behavioral regulation problems that have previously been identified in children with DCD are due to comorbidity with ADHD rather than DCD itself.
The current study has several distinct strengths, which include appropriate preprocessing techniques, and the use of a reliable and validated measure of behavioral regulation in a relatively large group that included children who met diagnostic criteria for ADHD, DCD and ADHD-DCD, as well as TD children. The measure used to assess behavioral regulation is well validated (Gioia et al., Reference Gioia, Isquith, Guy and Kenworthy2000) and although parent-reports are subjective, they capture a measure of behavior integrated over a longer time frame than can be observed in a laboratory visit and have better test−retest reliability (Enkavi et al., Reference Enkavi, Eisenberg, Bissett, Mazza, MacKinnon, Marsch and Poldrack2019). The study also has several weaknesses, including a relatively short scan time and differences between our groups of children with and without ADHD in (1) sex ratios, (2) IQ, and (3) distribution across scanners. We have done our best to account for these by including sex, IQ, and scanner as covariates in all analyses. While a short scan time is of benefit from an acquisition perspective, longer scan times may strengthen the reliability of FC estimates (Birn et al., Reference Birn, Molloy, Patriat, Parker, Meier, Kirk and Prabhakaran2013). It is also important to note that different task-based paradigms of behavioral regulation may yield additional insights to the resting-state paradigm employed here; we chose to investigate how an index score of behavioral regulation associates with FC across multiple brain networks to provide a more holistic perspective of the relationship between brain connectivity and behavioral regulation. Finally, while our FC maps were calculated using cross-correlation, a stronger measure than Pearson correlation, future work may be complemented by alternative FC measures that capture different aspects of FC (Mohanty et al., Reference Mohanty, Sethares, Nair and Prabhakaran2020).
Our findings significantly increase our knowledge on behavioral regulation and its underlying neural expression across a neurodiverse spectrum of children with and without ADHD, including children with DCD and combined ADHD-DCD. They suggest that behavioral regulation problems in DCD are likely attributable to comorbidity with ADHD. Children’s individual differences in behavioral regulation further associated with FC across diagnostic groups. Specifically, they associated with pathways between medial and lateral PFC, which may reflect a behavioral regulation process that integrates valuation and current behavioral goals. Children’s individual differences in behavioral regulation also associated with FC in frontostriatal reward pathways and visual reward pathways used in shifting of reward-directed action and inhibition of that action. Overall, our results highlight the utility of directly examining variables of potential clinical interest, such as behavioral regulation, and their associations with FC across children with differing neurodevelopmental conditions.
Acknowledgements
This work was supported by a grant from the Canadian Institutes of Health Research awarded to DD (MOP 88588), as well as NSERC CREATE I3T and Alberta Innovates Postdoctoral Fellowships awarded to CR. The funders had no role in study design, data collection and analysis, decision to publish or preparation of the manuscript. The authors thank all children and families who participated in this study.
Conflicts of interest
None.




