


Flavonoids are a large group of secondary plant metabolites with more than 6000 different compounds described to date. One important flavonoid subclass, the flavonols, and their major representative quercetin are ubiquitously distributed in plant foods and thus ingested in appreciable amounts with the daily diet. Particularly rich sources of quercetin are onions, apples and tea(Reference Hertog, Feskens and Hollman1, Reference Manach, Scalbert and Morand2). Epidemiological studies have revealed that the Mediterranean diet, which is particularly abundant in flavonoid-rich foods, correlates with increased longevity and a decreased incidence of CHD(Reference Williamson and Manach3, Reference Erdman, Balentine and Arab4). Dietary quercetin supplementation has recently been shown to lower platelet aggregation(Reference Hubbard, Wolffram and de Vos5) and blood pressure in man(Reference Edwards, Lyon and Litwin6). Furthermore, numerous cell culture studies indicate a potent antinflammatory activity of quercetin(Reference Lotito and Frei7–Reference Jung and Sung11).
ApoE is a polymorphic multifunctional protein with three common isoforms in man (E2, E3 and E4). ApoE3 is the wild-type and most common isoform, while apoE4 carriers account for about 25 % of the Caucasian population(Reference Eichner, Dunn and Perveen12). Presence of the apoE4 allele is associated with a 40–50 % higher risk of CVD(Reference Song, Stampfer and Liu13) and apoE4 is the major known genetic risk factor for maturity-onset Alzheimer's disease(Reference Mahley, Weisgraber and Huang14, Reference Cedazo-Minguez15).
Although apoE4 is strongly linked to both diseases, the molecular basis of these associations remains uncertain. Traditionally, the differences in disease risk have been attributed to the increased LDL-cholesterol (about 8 %) observed in apoE4 carriers, but it is becoming increasingly evident that this alone cannot explain the disease differential(Reference Davignon, Gregg and Sing16). ApoE is not only synthesised by the liver, but also in the brain and by resident macrophages(Reference Wilson, Myers and Larson17) in the atherosclerotic wall, where it exerts atheroprotective actions, independent of its role in lipid metabolism(Reference Miyata and Smith18).
Our group has previously shown that the apoE genotype significantly affects the cellular inflammatory response in stably transfected murine macrophages. In apoE4 v. apoE3 macrophages higher levels of proinflammatory cytokines including TNF-α were evident following stimulation with lipopolysaccharide(Reference Jofre-Monseny, Loboda and Wagner19). However, these findings refer to an in vitro situation and it is not known if the apoE genotype also affects biomarkers of inflammation in vivo. Furthermore, to the best of our knowledge no studies have examined the interaction between dietary flavonoids, apoE genotype and biomarkers of inflammation in vivo which is the aim of the present study.
Experimental methods
Mice and diets
Female homozygous apoE3 and apoE4 targeted replacement mice (6–8 weeks old) ‘humanized’ for the apoE gene were purchased from Taconic Europe (Ry, Denmark). Generation of the apoE3 and apoE4 models was recently described(Reference Jofre-Monseny, Huebbe and Stange20). Mice of both genotypes were assigned into one of two possible dietary groups (n 8), and were kept in macrolon cages at 21–25°C, with a 12 h day–night cycle.
The diets and water were provided ad libitum for 6 weeks and live weight recorded weekly. Mice were kept according to the German Regulations of Animal Welfare with permission of the responsible authority. The semi-synthetic diet, based on maize starch (15 %), casein (20 %), sucrose (33 %) and butter fat (21 %) (so-called ‘Western diet’), was purchased from Ssniff Special Diets (Soest, Germany). Diets were supplemented with 0 or 2 mg quercetin/g diet, as dihydrate ( ≥ 98 % purity; Roth, Karlsruhe, Germany).
Sample collection and tissue preparation
At the end of the dietary intervention period, mice were anaesthetised and decapitated. The blood was collected in EDTA-tubes, and the plasma was separated by centrifugation (8000 g, 4 min, 4°C) and stored at − 80°C. An aliquot of EDTA-blood was diluted 1:10 with RPMI 1640 medium (supplemented with 10 % fetal calf serum and 100 U/ml penicillin and 100 μg/ml streptomycin; all from PAA, Coelbe, Germany) and stimulated with 100 ng/ml lipopolysaccharide (from Salmonella enteritidis; Sigma, Deisenhofen, Germany) for 24 h as adapted from Miles et al. (Reference Miles, Zoubouli and Calder21). Accordingly, the supernatants were collected, centrifuged and cell-free samples were stored for further analysis at − 80°C. Mice were genotyped according to the method of Zivelin et al. (Reference Zivelin, Rosenberg and Peretz22).
Cholesterol levels and apoE concentrations
Plasma total cholesterol and apoE concentrations were determined on an ILAB 600 automatic analyser (Instrumentation Laboratories UK Ltd, Warrington, UK), using commercially available spectrophotometric kits (TC, Instrumentation Laboratories Ltd) and turbimetric immunoassay kits (Apolipoprotein E-HAWako; Alpha Laboratories Ltd, Eastleigh, UK).
TNF-α levels
TNF-α concentrations in the supernatants of whole-blood culture were determined by the mouse TNFα DuoSet ELISA development kit (R&D Systems, Wiesbaden, Germany).
Quercetin analysis
Analysis of plasma quercetin and its methylated derivative isorhamnetin (3′-O-methyl-quercetin) was performed by HPLC with fluorescence detection after treatment with a β-glucuronidase–sulphatase mix as previously described(Reference Ader, Wessmann and Wolffram23).
Statistical analysis
Statistical analysis was performed using SPSS version 13.0 (SPSS, Munich, Germany). Data were analysed for normality of distribution (Kolmogorow–Smirnov and Shapiro–Wilk tests) and equality of variance (Levene test) prior to two-way ANOVA, which was performed to test the independent effects of apoE genotype (apoE3 and apoE4), dietary quercetin levels (0 or 2 mg/g diet) and their interactions on the outcomes of interest. A t test was used to compare means of quercetin-fed mice with their control genotype. Data are expressed as means with their standard errors and significance was accepted at P < 0·05.
Results
Mice carrying the apoE3 and apoE4 genotype did not differ in live weight gain over the 6-week experimental period. Furthermore, there was no significant effect of dietary quercetin supplementation on final body weight in either the apoE3 or apoE4 transgenic mice (Table 1).
Table 1 Effect of apoE genotype and dietary quercetin on final body weight, plasma levels of quercetin, blood lipids and TNF-α levels in apoE3 and apoE4 targeted gene replacement mice‡
(Mean values with their standard errors)
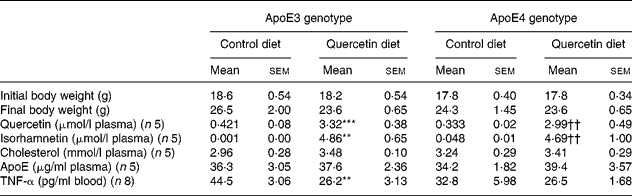
Mean values were significantly different from those of the apoE3 control group (t test): **P < 0·01, ***P < 0·001.
Mean values were significantly different from those of the apoE4 control group (t test): ††P < 0·01.
‡ For details of procedures, see Experimental methods. ApoE3 and ApoE4 targeted gene replacement mice were fed a diet without (control; 0 mg/g diet) or supplemented (quercetin diet; 2 mg/g diet) with quercetin for 6 weeks. Using two-way ANOVA, significant quercetin effects were observed for plasma quercetin (P < 0·001), isorhamnetin (P < 0·001) and TNF-α (P < 0·01).
Feeding the quercetin-supplemented diets significantly increased plasma levels of quercetin and isorhamnetin both in apoE3 and apoE4 mice to a similar extent. In mice fed the control diet quercetin was detectable in low concentrations whereas its metabolite isorhamnetin could only be determined in traces. There was no significant effect of apoE genotype on plasma quercetin and isorhamnetin levels (Table 1).
Table 1 shows mean values of plasma apoE and total cholesterol according to apoE genotype and dietary quercetin supplementation. ApoE3 and apoE4 transgenic mice exhibited similar plasma levels of both apoE and cholesterol which were not significantly affected by dietary quercetin. In mice receiving the basal diet without quercetin supplementation, levels of TNF-α in whole-blood supernatants after lipopolysaccharide stimulation were higher in apoE3 as compared to apoE4 transgenic mice. Dietary quercetin significantly lowered levels of TNF-α by 44 % in apoE3 mice (P = 0·0015). In apoE4 mice only a moderate decrease in TNF-α (20 %) due to dietary quercetin was evident. Following quercetin supplementation TNF-α levels were similar between apoE3 and apoE4 transgenic mice.
Discussion
Most studies in the literature reporting anti-inflammatory activities of quercetin were conducted in vitro using quercetin concentrations in the cell culture medium of 1–50 μmol/l(Reference Nair, Mahajan and Reynolds8, Reference Cho, Park and Kwon9). In the present study plasma levels of quercetin of 3·0–3·3 μmol/l were measured. Interestingly, this concentration was sufficient to induce a significant decrease in TNF-α levels after ex vivo stimulation of whole blood with lipopolysaccharide in apoE3 transgenic mice. The dietary quercetin level used in the present mouse study was set to be 2 mg/g diet, which refers to a quercetin concentration of approximately 400 mg/kg body weight. This is many times higher than quercetin concentrations which can be normally achieved due to a flavonoid-rich diet in man(Reference Manach, Williamson and Morand24). However, it needs to be taken into account that quercetin is also available in a purified form and commercially available quercetin supplements often contain quercetin concentration in a range between 500 and 1000 mg/capsule(Reference Weldin, Jack and Dugaw25).
In the present study a targeted gene replacement model was used to study interactions between apoE genotype and dietary quercetin. It is well documented that human apoE4 carriers exhibit lower levels of the apoE isoprotein as compared to man carrying the E3 allele(Reference Mahley and Rall26). Thus in man it is generally difficult to differentiate between genotype effects and effects which may be related to differences in the circulatory levels of the apoE isoprotein. Since our targeted gene replacement mice exhibited almost identical amounts of plasma levels of apoE3 and E4, respectively, potential differences in blood lipids and biomarkers of inflammation in the present mouse study are therefore related to the apoE3 and E4 isoprotein per se.
Human studies investigating the impact of apoE genotype on plasma lipid levels have reported mixed findings, with 5–10 % higher total- and LDL-cholesterol levels typically evident in E4 carriers(Reference Minihane, Jofre-Monseny and Olano-Martin27, Reference Minihane, Khan and Leigh-Firbank28), an effect which has been attributed to a number of mechanisms including increased dietary cholesterol absorption, decreased hepatic LDL receptor-mediated uptake and an increase in the production rate of LDL from its precursor VLDL. ApoE3 and apoE4 transgenic mice exhibited similar plasma levels of apoE and cholesterol which were not significantly affected by dietary quercetin supplementation. Cholesterol homeostasis and LDL receptor activity may also be affected by quercetin(Reference Pal, Ho and Santos29, Reference Davalos, Fernandez-Hernando and Cerrato30). However, in vivo effects of quercetin supplementation on human plasma lipid profile are inconsistent as well. No changes in plasma lipids were observed after quercetin supplementation in healthy subjects(Reference Chopra, Fitzsimons and Strain31, Reference Conquer, Maiani and Azzini32) whereas markedly lower blood lipids were found when quercetin (as grape powder/grape juice) was given to pre- and postmenopausal women(Reference Zern, Wood and Greene33) and to haemodialytic and healthy subjects(Reference Castilla, Echarri and Davalos34).
An important observation of the present study is that, although the differences did not reach statistical significance, apoE4 mice receiving the quercetin-free basal diet had 26 % lower levels of TNF-α as compared to apoE3 mice. However, the mouse data are in contradiction to our data in murine RAW264.7 macrophages where elevated levels of TNF-α were evident in E4 v. E3 macrophages(Reference Jofre-Monseny, Loboda and Wagner19). Thus cell culture data regarding apoE genotype and inflammation may not be directly extrapolated to an in vivo situation. Furthermore, it needs to be taken into account that apoE produced by macrophages contributes only to 20 % of the apoE produced in the human body whereas 80 % of apoE is produced in the liver. In accordance with the present TNF-α data we found also higher levels of C-reactive protein(Reference Jofre-Monseny, Huebbe and Stange20), another biomarker of inflammation, in apoE3 as compared to apoE4 transgenic mice.
Another finding of the present study is that dietary quercetin significantly decreased TNF-α levels in ex vivo-stimulated whole-blood samples of apoE3 but not apoE4 transgenic mice. Following quercetin supplementation in mice, TNF-α levels in apoE3 were lower than those in apoE4. Thus, based on the present findings, it seems likely that human apoE3 carriers may be more responsive towards a dietary intervention with flavonoids. Again, the present finding is in accordance with human data where supplementation with α-linolenic acid had a stronger effect on biomarkers of inflammation in apoE3 v. apoE4 carriers(Reference Paschos, Yiannakouris and Rallidis35). Therefore, the apoE genotype may in part explain the large heterogeneity of studies regarding potential health effects of flavonoids in man where cohorts are not genotyped for apoE polymorphisms.
The underlying molecular and cellular mechanisms by which dietary quercetin has resulted in a decrease of secreted TNF-α may be related to its ability to inhibit the NF-κB-dependent signal transduction cascades as reported earlier(Reference Nair, Mahajan and Reynolds8). Further studies are warranted to test the hypothesis whether NF-κB signalling in vivo is different in apoE3 v. apoE4 mice.
The present study reports, for the first time, that dietary quercetin differentially affects TNF-α levels in E3 and E4 targeted replacement mice. However, the study also has some limitations. First, the concentration of quercetin used in the present study was rather high; thus dose-dependent studies are warranted. Second, whole blood was stimulated with lipopolysaccharide to induce TNF-α secretion thus the inflammatory potential rather than the baseline level of circulatory TNF-α was determined. The model of ex vivo stimulation of whole blood has been previously used to evaluate the anti-inflammatory potential of dietary phenolics(Reference Miles, Zoubouli and Calder21). Finally, the duration of the experiment was set to be 6 weeks. Therefore it would be interesting to test the hypothesis whether chronic feeding with lower doses of quercetin over a longer period of time would affect TNF-α and other inflammatory cytokines in relation to apoE genotype. Furthermore, human studies are warranted to study interactions between dietary flavonoids, apoE genotype and biomarkers of chronic inflammation.
Acknowledgements
The project was supported by a grant of the German Ministry of Education and Science entitled ‘Functional Foods for Vascular Health’ (BMBF 0313856A). C. B.-S. and G. R. designed the study. C. B.-S., S. W. and A. M. M. performed the analyses. C. B.-S., A. M. M. and G. R. wrote, edited and reviewed the final manuscript. None of the authors have a known conflict of interest.

