


1. Introduction
Much of our understanding of global change is derived from studies of marine sediments, which have provided regionalto basin-scale records of environmental conditions such as ocean circulation and biological production. The Antarctic continental shelves are the most biologically productive regions of the Soulhern Ocean (Reference Comiso, McClain, Sullivan, Ryan and LeonardComiso and others, 1993), and are important areas of deep-water formation (Reference DeaconDeacon, 1984), yet palaeoenvironmental information for these waters is scant. This largely reflects the nature of marine sediments on the Antarctic continental margin: typically a complex mixture of poorly sorted terrigenous and biogenic material, often reworked by the action of ice and currents (Reference Anderson and MolniaAnderson and Molnia, 1989). However, some bathymétrie depressions such as relict Glaciol troughs and fjords may act as natural sediment traps, allowing the accumulation of relatively undisturbed sequences of marine sediments derived from overlying waters. in some cases, these deposits may provide detailed records of environmental conditions for these regions during the late Quaternary (see, e.g., Reference Domack, Mashiotta, Burkley, Ishman, Kennett and WarnkeDomack and others, 1993; Reference Leventer, Domack, Ishman, Brachfield, McClennen and ManleyLeventer and others, 1996). Because environmental conditions such as algal production and sea-ice cover are highly variable around the Antarctic continental margin (Comiso and others, 1993), numerous such sedimentary records are required to establish palaeoenvironmental records on a regional scale. The study described here aims to extract palaeoenvironmental information from sediments on the continental shelf off Mac. Robertson Land, East Antarctica (Mac. Robertson Shelf). This region sustains some of the highest phytoplank-ton biomass of the Antarctic continental margin (Comiso and others, 1993), and may be an area of significant bottom-water formation (NunesVaz and Lennon, 1996). We are using geochemical techniques to examine the accumulation of biogenic and lithogenic material in a scries of sediment cores recovered from the Mac. Robertson Shelf. Here we present geochemical data for two of these cores for which radiocarbon chronologies have been established, and discuss the palaeoenvironmental implications of these results.

Fig. 1. Map of the Mac. Robertson shelf, showing locations of sediment cores under study.
2. Methods
2.1. Sample materials
Sediment cores collected from the Mac. Robertson Shelf with RSV Aurora Australia in 1993,1995 and 1997 are used in the present study (Fig. 1). These 8 cm diameter gravity cores were collected from two deep areas of the shelf, the Nielsen Basin (>1000m water depth) and Iceberg Alley (<500m water depth), which are interpreted as relict Glaciol troughs (Harris and O'Brien, 1996). Results are presented here for two cores for which we have down-core radiocarbon dates and geochemical data, cores KROCK-GG1 (hereafter GC1) and KROCK-GC2 (hereafter GC2). GC1 was recovered from Iceberg Alley on the outer shelf, in 478 m of water, and GC2 was recovered from the Nielsen Basin on the inner shelf, in 1091 m of water. These sites are separated by a distance of roughly 90 km. Both cores recovered apparently continuous sequences of diatom ooze with minor amounts to line quartz, sand and other fine lithogenic material. GC1 has a total length of 363 cm ofolive-grccn ooze, with numerous ~l-10cm thick bands of fluffy lighter-coloured material, and ~5 cm of darker-coloured ooze at the base of the core. Core GC2 contains 293 cm of featureless olive-green ooze, except for ~10 cm of darker olive-grey sandy material at the base of the core. The cores were split immediately after collection, wrapped in polyethylene, and stored refrigerated to minimise water loss. The cores were subsampled during 1994-95 in 10 cm intervals for geochemical analysis. in addition, 1 cm subsamples were taken from the upper 10 cm of each core for determination of excess 210Pb; 1 -2 cm subsamples were taken at selected depths For radiocarbon dating; and 1cm3 subsamples were laken at 10 cm intervals for determination of dry bulk density. The uppermost section of GC1 shrank in length from 100 cm to approximately 90 cm prior to subsampling, presumably due to water loss. in the presentation of data for this core, average sample depths between 0 and 90 cm were multiplied by a factor of 100/90 in an effort to correct for shrinkage.
2.2. Core stratigraphy
The cores are essentially devoid of carbonate microfossils, precluding use of ![]() stratigraphy and radiocarbon dating of calcium carbonate. Measurements of uranium-series radionuclides (226Ra, 230Th, 231Pa) may allow estimates of accumulation rates, although dating techniques using these radioisotopes generally require uniform sedimentation rates, which may not (and probably do not) apply to these cores. The principal chronostratigraphic tool we have employed in this study is 14C-dating of bulk organic carbon. Radiocarbon dates for subsamples of these sediment cores, which contain around 1-2% organic carbon, were determined by accelerator mass spectrometry at the Australian Nuclear Science and Technology Organisation and at the New Zealand Institute of Geological and Nuclear Sciences. Radiocarbon dates are reported here as conventional radiocarbon years before present (BP), as defined by Reference Stuiver and PolachStuiver and Mach (1977). in addition, unsupported 2l0Pb was determined in the core-top subsamples by gamma spectrometry at the University of Hawaii, in an effort to evaluate recent accumulation rates, bioturbation and core-top losses.
stratigraphy and radiocarbon dating of calcium carbonate. Measurements of uranium-series radionuclides (226Ra, 230Th, 231Pa) may allow estimates of accumulation rates, although dating techniques using these radioisotopes generally require uniform sedimentation rates, which may not (and probably do not) apply to these cores. The principal chronostratigraphic tool we have employed in this study is 14C-dating of bulk organic carbon. Radiocarbon dates for subsamples of these sediment cores, which contain around 1-2% organic carbon, were determined by accelerator mass spectrometry at the Australian Nuclear Science and Technology Organisation and at the New Zealand Institute of Geological and Nuclear Sciences. Radiocarbon dates are reported here as conventional radiocarbon years before present (BP), as defined by Reference Stuiver and PolachStuiver and Mach (1977). in addition, unsupported 2l0Pb was determined in the core-top subsamples by gamma spectrometry at the University of Hawaii, in an effort to evaluate recent accumulation rates, bioturbation and core-top losses.
2.3. Geochemical measurements
The geochemical measurements performed on the core sub-samples include: bulk major and minor elements, determined in crushed, 60°C-dried (and, for major elements, deioniscd-water washed) portions of the 10 cm subsamples by X-ray fluorescence spectroscopy following a modification of the method of Reference ShimmieldShimmield (1984); biogenic silica (opal), determined in freeze-dricd portions of the 10 cm sub-samples by the method of Reference Mortlock and FroelichMortlock and Froelich (1989); and total organic carbon, determined in crushed, 60°C-dried, deionised-watcr washed portions of the 10 cm subsamples by the Isotope and Organic Geochemistry Laboratory of the Australian Geological Survey Organisation. The down-core geochemical data presented here are: (1) total Al and Fe, which provide an indication of terrigenous inputs if it is assumed that most of these elements are associated with terrigenous material (Reference Calvert and PedersenCalvert and Pederscn, 1993; Reference KumarKumar and others, 1995); (2) total Mn, which, when normalised to Al and compared with lithogenic abundance, provides information regarding down-core redox conditions (Calvert and Pederscn, 1993; Reference Yang, Elderfield, Pedersen and IvanovichYang and others, 1995); (3) total U and Mo, which, when normalised to Al and compared with lithogenic abundances, provide sensitive proxies of palaeo-rcdox conditions during sediment deposition (Calvert and Pedersen, 1993; Reference Crusius, Calvert, Pedersen and SageCrusius and others, 1996); (4) opal (assumed composition SiO2·0.4H2O), which has been used to infer palaeoproductivity in the Southern Ocean where the degree of post-depositional preservation is high or relatively constant (Reference Charles, Froelich, Zibello, Mortlock and MorleyCharles and others, 1991; Morllock and others, 1991); (5) total organic carbon (TOC), which has been used to infer palaeoproductivity in sediments from the Antarctic continental shelf (Leventer and others, 1996); and (6) excess (or biogenic) Ba, which in non-reducing sediments may provide a proxy record of productivity in overlying waters (Reference Dymond, Suess and LyleDymond and others, 1992). Biogenic Ba is calculated from total Ba minus lithogenic Ba, where lithogenic Ba is estimated as 0.0075A1 (0.0075 is the average crustal weight ratio of Ba/Al; Dymond and others, 1992).
The down-core geochemical data have been plotted against the average depth of each 10 cm subsample in the core.
3. Results and Discussion
3.1. Core preservation and chronostratigraphy
The down-core radiocarbon dates increase in a regular fashion for both cores (Fig. 2), suggesting that the cores contain continuous records of sedimentation extending back to the late Pleistocene for GC1 and mid-Holocene for GC2. The upper 10 cm of both cores contained no unsupported 210Pb, suggesting that at least ~200 years of the uppermost sediments have been lost during collection of the cores. This is not unexpected, given the relatively small bore of the gravity corer used and the high water content of the fine diatom ooze. Organic matter at the sediment-water interlace typically has a non-zero radiocarbon age, as a result of (1) the non-zero radiocarbon age of organic matter in the photic zone (assumed to be the principal source of organic carbon in these sediments), known as the reservoir effect, (2) bioturbation, and (3) dilution of fresh sediments by older, resuspended material, lb correct our raw radiocarbon ages (Fig. 2) for these processes, we have subtracted 1730 radiocarbon years BP, which is the measured age of a surface-sediment grab sample (AA186-GB9) recovered near the location of GC2 (this supplants an earlier correction of 1094 radiocarbon years BP used in Harris and others, in press). The corrected radiocarbon ages are presented in Figure 3, which we compare with the conventional radiocarbon time-scale. Using sedimentation rates of 82 cm ka−1 for the upper part of GC1, and 179 cm ka−1 for the upper part of GC2 (see section 3.3), the corrected core-top ages of 0.90 ka BP for GC1 and 0.30 ka BP for GC2 then suggest
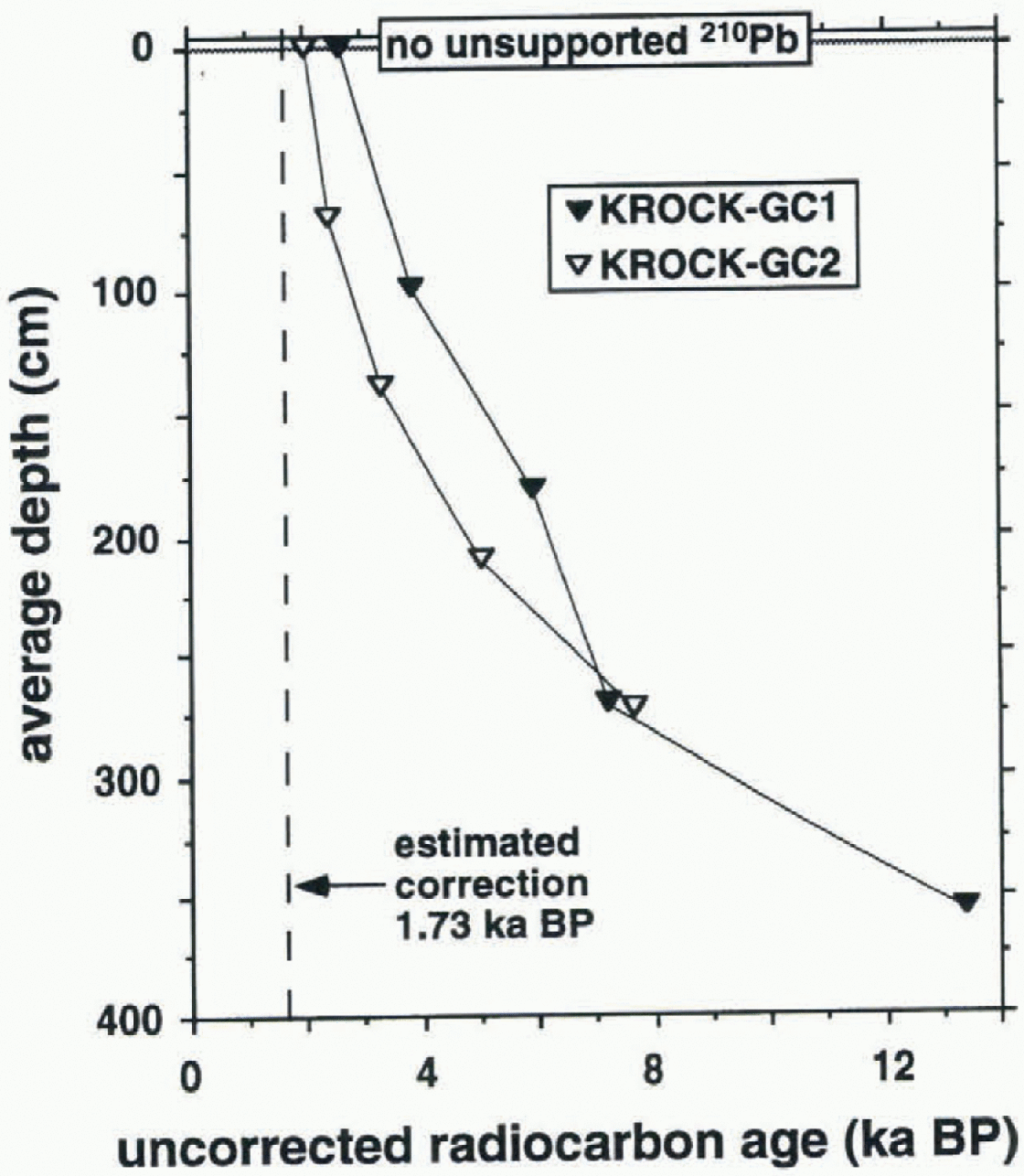
Fig. 2. Dawn-core uncorrected radiocarbon ages for cores KROCK-GC1 and KR0CK-GC2.
core-top losses of 74 and 54 cm, respectively. The radiocarbon data therefore suggest that GO represents a continuous sedimentary record from around 11700 to 900 radiocarbon years BP, and GC2 represents a continuous sedimentary-record from around 5940 to 300 radiocarbon years BP.
3.2. Down-core geochemical records
Down-core geochemical data (Al, Fe, Mn, Mo/Al, U/Al, opal, TOC and excess Ba) are presented for the two cores in Figure 3. Both cores are roughly similar in terms of bulk composition, containing around 30-50% opal and 0.5-2% TOC on a dry-weight basis. Down-core Mn concentrations are low and uniform in both cores, with Μn/Al close to the average shale ratio of 0.08 (Reference Wedepohl and AhrensWedepohl, 1971), whereas Mo and U are enriched relative to average shale ratios of Mo/Al ~ 0.3 x 10−4 and U/Al ~ 0.4 x 10 −4 (Wedepohl, 1971). Together, these trace-element data suggest that sediments accumulated at these sites under anoxic conditions, perhaps up to the sediment water interface (Calvert and Pedersen, 1993; Crusius and others, 1996). Such anoxic conditions promote diagenelic dissolution of biogenic barite and remobilisation of Ba with in the sediment column (Dymond and others, 1992), casting doubt on the utility of excess Ba as a palaeoproductivity proxy in these cores.
There are obvious contrasts in the down-core geochemical records of GC1 and GC2, suggesting significant differences in the composition, transport or preservation of sediments between the two sites. For GC1, there are four pronounced down-core minima in Al and Fe (shaded bands in fig. 3a) which are roughly coincident with local maxima in U/Al and Mo/Al (fig. 3b), and small local maxima in opal (fig. 3c) and TOC (fig. 3d). These down-core compositional changes suggest episodes of decreased terrigenous inputs (Al and Fe minima) and more reducing conditions with in the sediments (Mo and U maxima), associated with increased deposition of siliceous biogenic material (opal and TOC maxima). These compositional “episodes"

Fig. 3. Down-core geochemical data and corrected radiocarbon ages for cores KROCK-GC1 (a-e) and KR0CK-GC2 (f-j). Shaded bands indicate inferred bloom episodes.
coincide with layers of highest water content, and roughly correspond to some (but not all) of the thicker light-coloured bands in the core, except for the episode near the base of the core which occurs within relatively consolidated dark-coloured material.
We tentatively ascribe the down-core banding in GC1 to changes in species composition of diatoms with in the biogenic sediment fraction, rather than, for example, banding due to dilute sediment gravity flows. A micropalaeontologi-cal investigation of this core has shown that the diatom Fra-gilariopsis curia dominates the darker bands, whereas the lighter bands contain an increased abundance of the diatoms Core thron criophilum and Fragilariopsis cylindrus, and the darker material at the base of the core is characterised by an almost monospecific assemblage of Chaetoceros resting spores (personal communication from F. Taylor, 1997). Both C. criophilum and F. cylindrus are lightly silicified and thought to be poorly preserved in sediments (Jordan and others, 1991; Leventer and others, 1996; personal communication from F. Taylor, 1997), so the light-coloured bands in GC1 may result from massive blooms of these species in overlying waters. A simple interpretation of the down-core geochemical record for GO is that the minima in terrigenous components record episodes of enhanced export production by Corethron criophilum and/or Fragilariopsis cylindrus (or Chaetoceros at the base of the core), creating more reducing conditions with in the sediments. Calculated sediment accumulation rates for this core (see section 3.3) suggest that these compositional episodes, which we interpret as periods of enhanced export production, may have occurred over periods of ~100-1000 years, although these layers could have accumulated more rapidly over shorter periods. The numerous thin (~1-5cm) light-coloured bands with in the core may thus record shorter periods (< 100 years) of enhanced export production which are not discernible at the 10 cm resolution of our geochemical record.
It is interesting to note that the excess Ba concentrations calculated for GC1 are mostly less than zero (fig. 3e). This likely reflects (1) the diagenetic remobilisation of biogenic Ba under reducing conditions in the sediment column (thus rendering biogenic Ba unsuitable as a palaeoproductivity proxy), and (2) the presence of terrigenous components with a significantly lower Ba/Al ratio than the crustal average of 0.0075, such as quartz, in which A1 is known to substitute for Si (Reference Deer, Howie and ZussmanDeer and others, 1977). The lowest values of excess Ba calculated for GC1 roughly correspond to the upper three compositional episodes indicated by shaded bands in Figure 3, and may reflect increased diagenetic dissolution ofbiogenic baritc under relatively reducing sedimentary conditions accompanying enhanced export production. in this respect, calculated excess Ba may provide some qualitative palaeo-environmental information in anoxic continental-shelf sediments, with low concentrations indicating more reducing conditions with in the sediment column.
The down-core chemical changes observed in GC1 are generally absent from GC2, except at the base of the core which grades into sandier material. This core contains slightly lower concentrations of A1, Fe and Mn (fig. 3f), significantly higher ratios of Μο/Al and U/Al (fig. 3g), slightly higher concentrations of opal (fig. 3h) and significantly higher concentrations of TOC (fig. 3i) than GC1. Down-core diatom assemblages are dominated by Fragilariopsis curia (personal communication from F. laylor, 1997). These data suggest that the accumulation ofbiogenic and terrigenous material at the site of GG2 has been relatively consistent since the mid-Holocene, with a greater proportion of biogenic material and thus a more reducing sedimentary environment than at the GC1 site. However, some down-core geochemical variations are observed for GC2: changes in U/Al and particularly Mo/Al (fig. 3g), which suggest temporal variations in the sedimentary redox environment, and a general decrease in TOC with depth (fig. 3i), which may reflect either a decreased accumulation of organic material in older sediments, or the diagenetic decomposition of organic carbon with increased depth in the sediment column (Reference BernerBerner, 1980). Another contrast with GC1 is the calculated excess Ba concentrations in GC2, which are relatively uniform and all greater than zero (fig. 3j). This may reflect a higher Ba/Al ratio in the lithogenic material in GC2, or simply a greater deposition of biogenic barite such that accumulation of biogenic Ba has exceeded diagenetic loss from the sediment column.
3.3. Sediment accumulation rates
The down-core radiocarbon dates allow the calculation of average sediment accumulation rates between the dated sample depths for the two cores, which are shown in figure 4a and d (for GC1 and GC2, respectively). Compaction has been neglected in these calculations, and is assumed to be small. If compaction is defined as water loss due to compression from overlying sediment (Berner, 1980), then this assumption is probably valid: large down-core variations in water content in GC1 (~40-90% waler; data not shown) primarily reflect the observed layering with in the core, and show no systematic decrease with depth, whereas for GC2 the decrease in down-core water content is less than 10% with in the diatom ooze facies (~70-80% water; data not shown). Together with down-core bulk concentrations and measurements of dry bulk density, these calculated sediment accumulation rates may be used to estimate down-core material accumulation rates. These rates are presented for biogenic components (opal and TOC) in figure 4b and e (for GC1 and GC2, respectively), and for lithogenic components (Al and Fe) in figure 4c and f (for GC1 and GC2, respectively).

Fig. 4. Down-core corrected radiocarbon ages, average accumulation rates, and mass accumulation rates of opal, TOC, Al and Fe for cores KR0CK-GC1 (a-c) and KR0CK-GC2 (d-f).
The average accumulation rates calculated for GC1 for the mid- to late Holocene range from 39 to 82 cm ka−1 (fig. 4a). However, it should be noted that there are considerable uncertainties in these values arising from the observed layering in this core, which may represent short periods of rapid sediment accumulation that are unrcsolvable using radiocarbon dating. The average accumulation rate of 13.7 cm ka−1 calculated for GC1 in the early Holocene is likely too low, because it uses the 11660ka radiocarbon age obtained for darker material at the base of the core, which may include slowly accumulating and/or reworked pre-Holocene sediment. A more accurate estimate to sediment accumulation rale for this site during the early Holocene will require a radiocarbon date with in the early Holocene (<10 ka BP) section of the core (around 300-350 cm depth). The down-core material accumulation-rate estimates vary widely for GO, and these variations primarily reflect the large changes in down-core dry bulk density (0.08-0.9 g cm−3) and the low accumulation rate calculated for the lower section of the core. We believe that the accumulation-rate estimates shown in figure 4b and c are not representative of actual material accumulation rates for GO, because (1) the low values of dry bulk density reflect the fluffy layers which may have accumulated faster than the calculated average accumulation rates, and (2) the average accumulation rate calculated for the lower section of core is likely too low, as discussed above.
The average accumulation rates and material accumulation rates calculated for GC2 (fig. 4d-f) are probably more reliable than those calculated for GO, because all radiocarbon dates are for material with in the Holocene facies, and down-core bulk densities are relatively uniform (0.27 ± 0.05 g cm −3). The calculated average accumulation rates for GC2 suggest a dramatic (<7-fold) increase in sediment accumulation rates from the middle to late Holocene, from 24.5 cm ka−1 between 3330 and 5940 radiocarbon years BP to 179 cm ka−1 between 300 and 690 radiocarbon years BP (fig. 4d). Further, the estimated material accumulation rates suggest that there has been a similar 5-10-fold increase in the accumulation of biogenic (opal and TOG) and lithogenic (AÍ and Fe) material at this site from the middle to late Holocene (fig. 4e and f).
Two general hypotheses may be offered to expla in the apparently synchronous increases in the accumulation rate ofbiogenic and lithogenic material at the location of GC2 during the Holocene. The first is that the apparent increase in sediment accumulation rate is the result of changes in sediment transport processes or sediment focusing, such that sediments were transported into the inner Nielsen Basin from a progressively increasing “catchment” area during the Holocene, due to retreat of the ice sheet from the shelf or changes in shelf-water circulation. The second hypothesis is that the apparent increase in sediment accumulation rates reflects real increases in pelagic sedimentation in overh ing waters, perhaps as a result of progressively decreasing permanent ice cover at this site during the Holocene, allowing comparable increases in biogenic export production and deposition of ice- and current-borne lithogenic material. Another possibility consistent with the second hypothesis is that primary production in these shelfwaters is limited by the availability of dissolved iron in surface sea water, as has been suggested for offshore Antarctic waters (Reference Martin, Fitzwater and GordonMartin and others, 1990) and shelfwaters of the Ross Sea (Scdwick and DiTullio, 1997). in this case, the release of iron from lithogenic material entering surface waters might regulate algal export production.
4. Conclusions and Future Research
The results so far obtained for cores KROCK-GCl and KROCK-GC2 suggest quite different sedimentation histories for these two shelf locations during the Holocene. On the outer shelf, GO apparently records three episodes of intense export production during the Holocene separated by-periods of around 1500 years, and probably less pronounced “bloom episodes” which occurred over shorter periods. Long-term average sediment accumulation rates range from 39 to 82 cm ka−1 during the middle and late Holocene. At present, our palaeoenvironmental interpretation of this core is limited by the 10 cm (>100year) resolution of the geochemical analyses and the much coarser resolution of the radiocarbon dates. Higher-resolution stratigraphie, geochemical and sedimentological analyses of this or similar cores, ideally at the scale of the visible bands with in the core (i.e. t in the order of 1-10 cm), will allow us to better constra in the timing and duration of these bloom episodes and to better quantify material accumulation rates.
In contrast, our analysis of GC2 from the inner shelf suggests that there has been a roughly constant proportion of biogenic and lithogenic material accumulating at this location during the middle to late Holocene, with a somewhat higher proportion of biogenic material relative to the outer shelf. Notably, there is an apparent 7-fold increase in average sediment accumulation rate (from 24.5 to 179 cm ka−1) at this location between the middle and lale Holocene, with roughly comparable increases in the accumulation of both biogenic and lithogenic material. This may represent changes in sediment transport processes; i.e. sediment focusing, or real increases in pelagic sedimentation at this location during the Holocene. in an effort to distinguish between these two possibilities, we are currently measuring 230Th and 23lPa in GC2, which offers the possibility of quantifying sediment focusing (Reference FrankFrank and others, 1995) and evaluating changes in biogenic export production (Kumar and others, 1995).
Acknowledgements
The authors gratefully acknowledge P. O'Brien for collection of core material and initial encouragement, R. Connell for laboratory assistance, ETaylor for diatom identification, and T. Trull, E. Domack, I. Goodwin and R. Anderson for insightful discussions. E. Domack and J. Anderson are thanked for reviewing the manuscript. Staff of the Isotope and Organic Geochemistry Laboratory of the Australian Geological Survey Organisation are thanked for performing the TOC determinations. Uranium-series measurements were accomplished with travel funding provided by the Australian Departmenl of Industry, Science and Technology. The staff of the Australian Nuclear Science and Technology Organisai ion accelerator mass spectrometry facility are thanked for radiocarbon analyses, which were funded by grants from the Australian Institute of Nuclear Science and Engineering. The principal funding for this work was provided by the Antarctic CRC.




