



The seminal idea of animacy effect was introduced by Nairne, VanArsdall, Pandeirada, Cogdill, and LeBreton (Reference Nairne, VanArsdall, Pandeirada, Cogdill and LeBreton2013) in their study of the mnemonic value of animacy. Bonin, Gelin, and Bugaiska (Reference Bonin, Gelin and Bugaiska2014) have provided a definition of animacy effect: “Animacy effect refers to the observation that animate stimuli are remembered better than inanimate stimuli” (p. 371). A number of studies have investigated the animacy effect extensively, and found it is pervasive and robust. By counterbalancing factors such as familiarity and word frequency, which may potentially influence memory, animacy effect is still observed in free recall tasks (Nairne et al., Reference Nairne, VanArsdall, Pandeirada, Cogdill and LeBreton2013), recognition tasks (Bonin et al., Reference Bonin, Gelin and Bugaiska2014; Gelin, Bugaiska, Meot, & Bonin, Reference Gelin, Bonin, Meot and Bugaiska2017), and paired-associate learning tasks (VanArsdall, Nairne, Pandeirada, & Cogdill, Reference VanArsdall, Nairne, Pandeirada and Cogdill2015). With regard to the types of material, animacy effect is found in studies that use word and picture stimuli (Bonin et al., Reference Bonin, Gelin and Bugaiska2014), and even non-words contaminated with properties of living things (VanArsdall, Nairne, Pandeirada, & Blunt, Reference VanArsdall, Nairne, Pandeirada and Blunt2013).
An ultimate explanation of the animacy effect proposed by Nairne et al. (Reference Nairne, VanArsdall, Pandeirada, Cogdill and LeBreton2013) is that the human memory system is tuned to solve adaptive problems. As animals have represented both prey and predators in the history of human evolution, they have played an important role in human survival and reproduction. Thus, humans must be able to rapidly recognize animals and preserve them in long-term memory (Nairne, Reference Nairne2010).
Though this explanation is powerful, researchers have devoted themselves to the underlying proximate mechanisms of animacy effect, which have not yet been determined. However, studies have failed to support the fact that well-established cognitive mechanisms are responsible for the animacy effect. For example, it was found that sensory feature (Bonin et al., Reference Bonin, Gelin and Bugaiska2014), intention and memory load (Bonin, Gelin, Laroche, Meot, & Bugaiska, Reference Bonin, Gelin, Laroche, Meot and Bugaiska2015), encoding context (Gelin et al., Reference Gelin, Bugaiska, Meot and Bonin2017), mental arousal (Popp & Serra, Reference Popp and Serra2017), categorical recall strategy (VanArsdall, Nairne, Pandeirada, & Cogdill, Reference VanArsdall, Nairne, Pandeirada and Cogdill2017), levels of processing (Leding, Reference Leding2018), imagery processes (Gelin, Bugaiska, Meot, Vinter, & Bonin, Reference Gelin, Bugaiska, Meot, Vinter and Bonin2018), and emotional arousal (Meinhardt, Bell, Buchner, & Roer, Reference Meinhardt, Bell, Buchner and Roer2018) are not the reasons for animacy effect. Thus, it is still unclear that the benefit of memory of animate stimuli is due to any domain-general cognitive mechanism that is responsible for the animacy effect.
The animacy effect refers to animate stimuli that are more likely to be retrieved preferentially compared to other stimuli. The successful retrieval of targets in memory does not only depend on the activation of targets, which must be sufficient to reach the threshold for recall (Collins & Loftus, Reference Collins and Loftus1975), but on the inhibition of irrelevant information (Anderson & Green, Reference Anderson and Green2001). Because irrelevant information competes with target information during the retrieval process, a mechanism of inhibiting irrelevant information is triggered to diminish the competition with target information. The suppression of irrelevant information is not a passive side effect of storing new memories, but results from inhibitory control mechanisms recruited to override the unwanted memories (Anderson, Reference Anderson2003). Therefore, the inhibitory process may underlie the control of memory (Schilling, Storm, & Anderson, Reference Schilling, Storm and Anderson2014) and reflect the action of a general executive control process (Anderson & Green, Reference Anderson and Green2001) that should be responsible for the better memory of animate stimuli.
In view of the inhibitory control theory, the memory advantage of certain targets may depend on the ability to counteract inhibitory control. Though animate stimuli have not been investigated, studies conducted with stimuli that are important to human survival have revealed the insensitivity to the inhibitory control. For example, stimuli that elicit negative emotions, such as threat and anxiety, have been shown to reduce or eliminate the effects of inhibition (Giebl, Storm, Buchli, Bjork, & Bjork, Reference Giebl, Storm, Buchli, Bjork and Bjork2016; Kircanski, Johnson, Mateen, Bjork, & Gotlib, Reference Kircanski, Johnson, Mateen, Bjork and Gotlib2016; Kuhbandner, Bäuml, & Stiedl, Reference Kuhbandner, Bäuml and Stiedl2009). These studies imply that animate stimuli may be endowed with the ability to counteract the inhibitory control. The aim of this study is to find out the role of inhibitory control in animacy effect. We have found a procedure called the retrieval practice paradigm, which is believed to be an effective tool for examining inhibitory control (Noreen & MacLeod, Reference Noreen and MacLeod2015).
Anderson, Bjork, and Bjork (Reference Anderson, Bjork and Bjork1994) introduced the retrieval practice paradigm in their pioneering study of retrieval-induced-forgetting (RIF). The RIF is a phenomenon in which the retrieval of items from a category impairs the memory of the other items in the same category. In standard RIF experiments, subjects are exposed to items from various categories presented in the form of “category-exemplar” (e.g., fruit-orange) in the learning phase. In the subsequent retrieval practice (RP) stage, only half of the items in a category are practiced. Subjects are cued with “fruit-or__” and are asked to complete the missing letters. Items for retrieval practice are marked as RP+, and the remaining items in the same category are marked as RP-. Items in other categories are marked as NRP. At the final test phase, subjects are asked to recall all the items they have learned. By comparing RP- items to NRP items, researchers have found that memory of RP- items is impaired. Anderson et al. (Reference Anderson, Bjork and Bjork1994) provide an inhibitory account of RIF and believe it is the consequence of inhibitory control of RP- items (Anderson & Spellman, Reference Anderson and Spellman1995). According to the inhibition-based theory, impairment of RP- items results from competitive memories that are activated by retrieval cues. To facilitate the retrieval of targets (RP+) during the retrieval practice phase, an inhibitory control is actively triggered to diminish competition from unpracticed items (RP-). Thus, the active inhibition is the mechanism underlying forgetting.
In this study, the inhibitory control on both animate and inanimate stimuli have been manipulated to test the hypothesis that animacy effect is due to the insensitivity of animate stimuli to the inhibitory control. Aslan and Bauml (Reference Aslan and Bauml2010, Reference Aslan and Bauml2012) suggest that the elimination of RIF results from the lack of inhibitory processing. Therefore, if the animacy effect is induced by the insensitivity to inhibition, the recall of inhibited animate items should be equal to those are uninhibited. In contrast, the inanimate items should be significantly influenced by the inhibitory control. Given the absence of inhibitory effect is the direct evidence to support the assumption of this study, the data are more likely under the null model compared to the alternative model. We have complemented the null significant hypothesis test (NSHT) with a corresponding Bayes factor hypothesis test, which allowed us to quantify evidence for the null hypothesis H0 (Hu, Kong, Wagenmakers, Ly, & Peng, Reference Hu, Kong, Wagenmakers, Ly and Peng2018; Wagenmakers et al., Reference Wagenmakers, Marsman, Jamil, Ly, Verhagen, Love and Morey2018).
EXPERIMENT 1
In experiment 1, we manipulated the inhibitory control into two semantic categories (animal and artifact) by using the retrieval practice paradigm in which the impaired memory is believed to represent the result of inhibition. Besides examining the role of inhibitory control in the memory of the two categories, we expected to replicate the animacy effect.
Different from the standard RIF paradigm, we measured the inhibitory effect by comparing the RP- items to the NRP items in the same category instead of between categories. This change is based on the assumption that the memory performance on every category would be equal without any experimental treatment if the inhibitory effect was measured by the comparison of RP- items and NRP items between categories. However, VanArsdall et al. (Reference VanArsdall, Nairne, Pandeirada and Cogdill2015) suggest that animacy should be considered as an extra mnemonic dimension to be controlled in memory research concerned with animate materials. It is inappropriate to compare animate items to other items directly, because animate stimuli tend to be remembered much better.
Method
Participants
One hundred and ninety-two Harbin University undergraduates (86 males) with an average age of 19.66 ± 1.40 years were recruited and instructed to complete an experiment of memory and reasoning. Participants were randomly divided into three groups (64 in each group). The first group was a control group in which participants were excluded from the inhibitory control. The other two groups were the experimental groups in which participants were exposed to the inhibitory control to animate stimuli or inanimate stimuli respectively. All the participants reported normal or corrected-to-normal visual acuity and were not told the purpose of the experiment. Gifts were provided to all the participants as a reward.
Materials
As the memory advantage for animate stimuli is the result of evolution, we selected artifacts as a control condition to compare the effects of inhibition. Many researchers use artifacts as materials for control conditions in studies of the animacy effect (Gelin, Bonin, Meot, & Bugaiska, Reference Gelin, Bonin, Meot and Bugaiska2017; Gelin et al., Reference Gelin, Bugaiska, Meot and Bonin2017; Li, Jia, Li, & Li, Reference Li, Jia, Li and Li2016; Popp & Serra, Reference Popp and Serra2016, Reference Popp and Serra2017; VanArsdall et al., Reference VanArsdall, Nairne, Pandeirada and Cogdill2017). Because artifacts have appeared more recently in human history than animals, the domain-specific cognitive processing have not yet evolved for these stimuli. Therefore, artifacts are well suited for distinguishing the mental processes underlying animate stimuli.
Materials were selected from the database of Liu, Hao, Li, and Shu (Reference Liu, Hao, Li and Shu2011). Sixty subjects re-rated these words to produce norms for familiarity (from 1 for the most unfamiliar to 5 for the most familiar) and subject word frequency (from 1 for the lowest to 7 for the highest frequency). Thirty-two words (16 animals and 16 artifacts) were selected by carefully matching both familiarity and subject word frequency between categories. The t-test results indicated no significant difference between the two categories on familiarity (animal: M = 4.70, SD = 0.16; artifact: M = 4.73, SD = 0.14), t(30) = −0.66, p > .05, d = −0.23, and subject word frequency (animal: M = 2.29, SD = 0.31; artifact: M = 2.16, SD = 0.40), t(30) = 1.02, p > .05, d = 0.36. To ensure the versatility of the materials, we adopted “morphological productivity” as an index to illustrate the effectiveness of chosen materials. Morphological productivity is defined as the ability of a Chinese character to produce words and is measured by the number of words that began with the character in this study. The data were obtained from a website (zuci.51240.com) that provides a service to list all the words containing the testing character. We only selected the words that began with the testing character. The t-test results indicated no significant difference between the two categories on the dimension of morphological productivity (animal: M = 437.69, SD = 549.97; artifact: M = 245.63, SD = 59.24), t(30) = 1.28, p > .05, d = 0.45. Each word was composed of two Chinese characters. The first and second characters were varied across items (see Appendix).
Procedure
Participants were tested up to four at a time in a quiet room for approximately 35 minutes. In the learning phase, 32 stimuli were presented on the computer screen successively in the form of category-exemplar pairs, such as “动物-蝴蝶” (animal-butterfly) and “人造物-信封” (artifact-envelope). The participants were required to memorize both animal and artifact items. Each category-exemplar pair was presented for 5 seconds on the screen. The sequence was counterbalanced between participants in order to exclude the serial position effect.
In the subsequent retrieval practice phase, half the items of a category were selected for retrieval practice (RP+ items). Each RP+ item was retrieval practiced three times in a random order across participants. Participants were cued every 10 seconds by the first character together with its category name (e.g., 动物-蝴__), which were presented in a booklet (one cue per page). They were asked to complete the items with the word they recollected from memory and write the missing character on the booklet. There was a reminder sound every 10 seconds to prompt turning to the next page. Participants in the condition of inhibitory control to animals were retrieval practiced with animate items, while participants in the condition of inhibitory control to artifacts were retrieval practiced with inanimate items. There were 32 sets of RP+ items (16 animate sets and 16 inanimate sets) in which the RP+ items were different from one another. The control group was excluded from the retrieval practice phase.
In the distracting phase, participants in the three groups were asked to complete as many Sudoku tasks as possible within 15 minutes. The Sudoku tasks were arranged in a sequence from very easy to very hard. In the final recall testing phase, participants were required to recall all the items they had learned in the learning phase, and write the items on an answer sheet within 5 minutes, without considering the categories or sequence of items from the learning phase.
Data analysis
The animacy effect was measured by the comparison of NRP items between two categories, while the inhibitory effect was measured by the comparison between the RP- and NRP items within category. The criterion for judging the presence of inhibitory effect was a significantly lower recall of the RP- items than that of the NRP items. The RP+ items were excluded from the data analysis as they were irrelevant to the measurements of animacy effect and inhibitory effect. The data of NRP items were obtained from the control group, and the data of RP- items were collected from the two experimental groups. Therefore, we employed an independent samples t test in favor of the measurements of animacy effect and the inhibitory effect.
The recall ratio for RP- items was calculated by the number of RP- items correctly recalled, divided by the number of all RP- items (8 items were included in the present experiment). The recall ratio for NRP items was calculated by the number of items correctly recalled from the uninhibited category divided by the total number of items in the category (16 items were included in the present experiment).
The Bayes factor hypothesis test were conducted using JASP software (https://jasp-stats.org/; JASP Team, 2018) with default prior width (Cauchy scale = 0.707). The BF01 was reported in this study because it is more convenient to interpret the absence of inhibitory effect compared to BF10 (Hu et al., Reference Hu, Kong, Wagenmakers, Ly and Peng2018). For example, BF01 = 10 means that the observed data is 10 times more likely to support the null hypothesis H0 than the alternative hypothesis H1.
Results
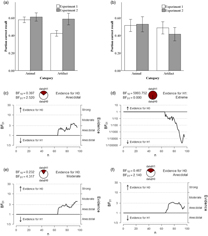
The descriptive statistic results are presented in Table 1. The results indicated animals (M = 0.366, SD = 0.133) were remembered better than artifacts (M = 0.266, SD = 0.115), t(126) = 4.590, p < .001, d = 0.811, suggesting a strong animacy effect was observed (see Figure 1a). The result of Bayes factor analysis also indicated an extremely strong evidence to support the presence of animacy effect, BF01 = 0.001 (see Figure 1b). A strong effect of inhibition on animals was detected (RP-: M = 0.268, SD = 0.186; NRP: M = 0.366, SD = 0.133), t(126) = −3.452, p < .001, d = −0.610, but no effect was observed on artifacts (RP-: M = 0.252, SD = 0.172; NRP: M = 0.266, SD = 0.115), t(126) = −0.530, p > .05, d = −0.094. The Bayes factor analysis revealed the similar results (inhibition to animals: BF01 = 0.028; inhibition to artifacts: BF01 = 4.662, see Figure 1c and 1d). The raw data are provided in the supplementary material.
Table 1. The descriptive statistic of the portion of correct recall in Experiment 1
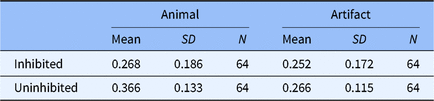
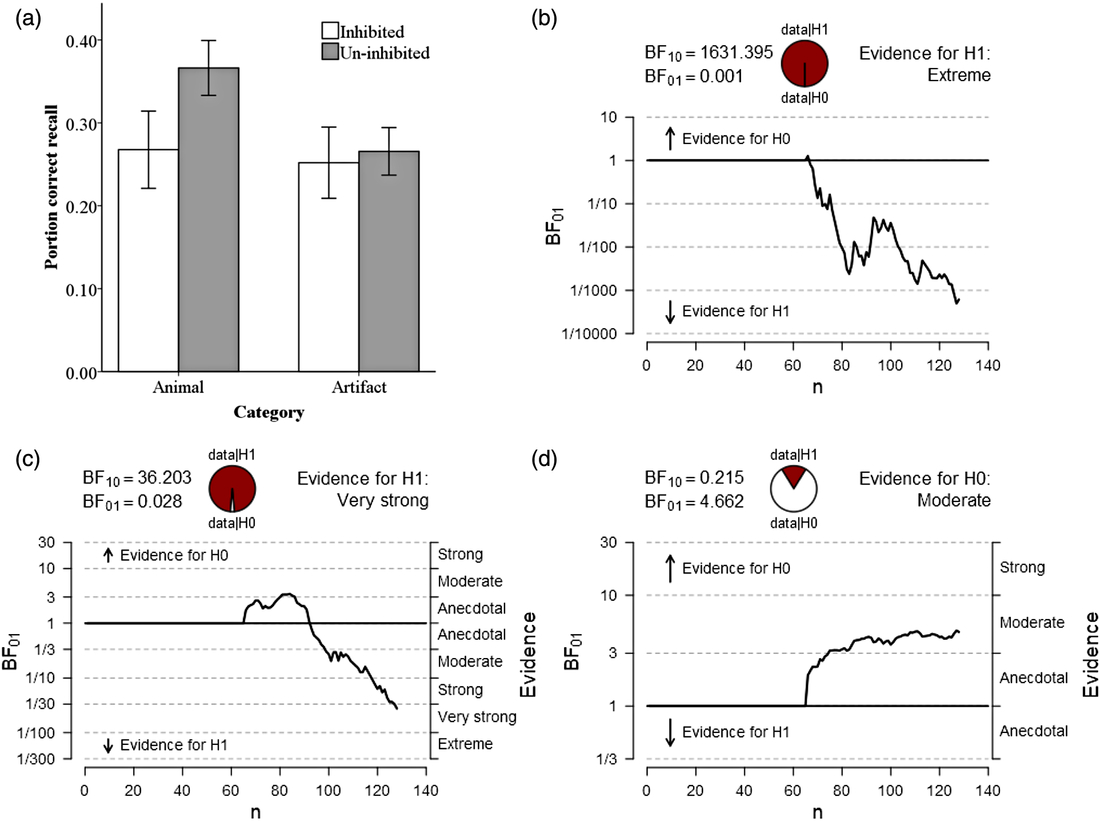
Fig. 1. Results from Experiment 1 analyzed using: (a) null significance hypothesis testing (error bars denote ± 2 SE) and sequential analysis from Bayesian independent t tests for (b) the animacy effect compared by NRP items between animal and artifact; (c) inhibitory effect on animals and (d) inhibitory effect on artifacts.
Discussion
Consistent with the previous studies, the results revealed a strong animacy effect in this study. We were frustrated to observe that the memory of RP- items of animal was impaired by the retrieval practice, suggesting the inhibitory control had significantly influenced the memory of animate items. The evidence did not fit the hypothesis that animacy effect was the result of the insensitivity to the inhibitory control.
Unexpectedly, the results indicated the absence of inhibitory control to artifacts, suggesting that memory of artifact was not influenced by the inhibitory process as the inhibitory account of RIF predicted. The memory of the NRP items of artifacts was little, as it was inhibited by the retrieval practice, despite the fact that the data of NRP items and RP- items were collected from two independent groups. If the inhibitory account for the RIF failed to explain the results of retrieval practice paradigm, the absence of inhibitory effect would also been found in the animate category. There would be an extra mechanism attenuating the memory of NRP items of artifact.
According to the inhibitory control account of retrieval (Anderson & Green, Reference Anderson and Green2001; Bjork, Reference Bjork, Roediger and Craik1989; Levy & Anderson, Reference Levy and Anderson2002), the retrieval process is a kind of selection-plus-suppression mechanism. It is argued that inhibitory resources are recruited to prevent unwanted or irrelevant memories from coming to mind, thereby promoting the retrieval of targets. Thus, the effect of inhibition on artifacts may be moderated by the competition between animals and artifacts during free recall phase. This means the artifacts are more likely to be inhibited by the memory of animals. With the paradigm used in Experiment 1, both animals and artifacts are memorized within a memory set. The animals may be selected as targets, owing to the advantage of memory, and the inhibitory control is activated to suppress the competition of artifacts. Therefore, the artifacts may be inhibited by the inhibitory control mechanism to facilitate the retrieval of animals. The second experiment was conducted to exclude the influence of the potential inhibition induced by the memory of animals.
EXPERIMENT 2
In Experiment 2, we excluded the potential influence of the inhibition resulting from the memory of animals by manipulating the animacy as a between-subject variable instead of a within-subject variable as it was in Experiment 1. In doing this, we expected a resurgence of the inhibitory effect on inanimate items.
More importantly, the absence of animacy effect was predicted in Experiment 2. It should be noted, however, that all the previous studies reporting the presence of animacy effect have employed experiments with animacy designed as a within-subject variable. As stated in the discussion part of Experiment 1, the tendency to retrieve animate items would suppress the inanimate items in the same memory set. Once removed from the inhibition of a competitive category, such as animals, inanimate items would be recalled as equal as animate items.
Method
Participants
One hundred and twenty-eight Harbin University undergraduates (54 males) with an average age of 18.39±1.13 years were recruited and asked to complete an experiment of memory and reasoning. All the participants reported normal or corrected-to-normal visual acuity and were not informed of the purpose of the experiment. Gifts were provided to all the participants as rewards.
Materials
Materials were identical to Experiment 1.
Procedure
Participants were tested up to four at a time in a quiet room, and were randomly divided into four groups (32 in each group). Two of the four groups were tested with either animals or artifacts without retrieval practice, while the other two groups were tested with either animals or artifacts items with retrieval practice. Thus, the NRP of animals or artifacts were acquired from the first two groups, and RP- of animals or artifacts were acquired from the other two groups. Besides the presenting stimuli of 16 animals or artifacts, the procedure was identical to Experiment 1. The measurements of animacy effect and inhibitory effect were the same as Experiment 1.
Results
Descriptive statistic results are presented in Table 2. No animacy effect was observed according to the results of Experiment 2. Animals (M = 0.609, SD = 0.133) were remembered as equally as artifacts (M = 0.586, SD = 0.186), t(62) = 0.579, p > .05, d = 0.145 (see Figure 2a). The result of Bayes factor analysis also indicated moderate evidence to support the absence of animacy effect, BF01 = 3.395 (see Figure 2b).
Table 2. The descriptive statistic of the portion of correct recall in Experiment 2
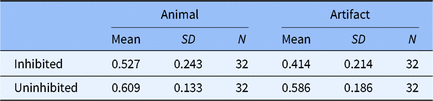
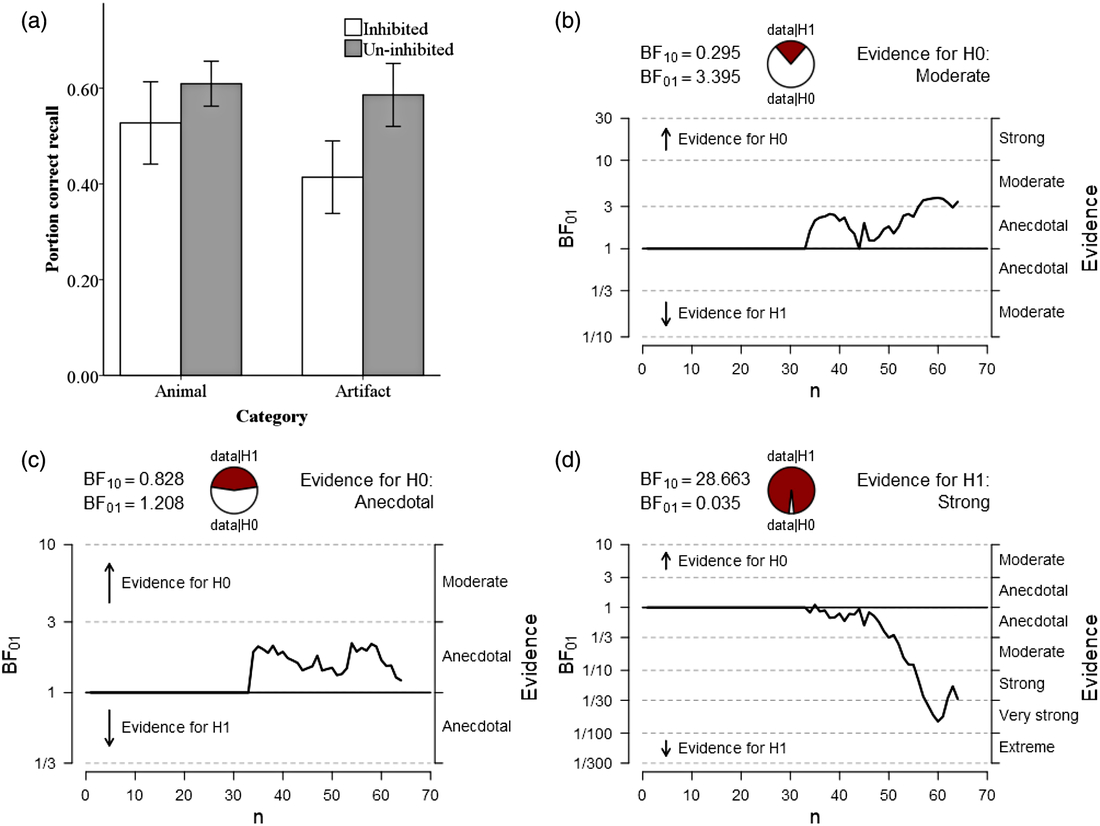
Fig. 2. Results from Experiment 2 analyzed using: (a) null significance hypothesis testing (error bars denote ± 2 SE) and sequential analysis from Bayesian independent t tests for (b) the animacy effect compared by NRP items between animal and artifact; (c) inhibitory effect on animals and (d) inhibitory effect on artifacts.
The results indicated the absence of inhibitory effect on animals (RP-: M = 0.527, SD = 0.243; NRP: M = 0.609, SD = 0.133), t(62) = −1.674, p > .05, d = −0.418, but strong effect on artifacts (RP-: M = 0.414, SD = 0.214; NRP: M = 0.586, SD = 0.186), t(62) = −3.426, p < .01, d = −0.857. The Bayes factor analysis revealed the effect of inhibition on animals was anecdotal, BF01 = 1.208 (see Figure 2c), but strong to support the presence of inhibitory effect on artifacts, BF01 = 0.035 (see Figure 2d).
In order to ensure that the absence of animacy effect was due to the lack of inhibition resulting from the memory of animals, we compared the results of the two experiments. However, the list lengths of the two experiments were not equal, and the list length effect (Ward, Reference Ward2002), which predicts that the number of recalls increases but the rate of recalls decreases as the list length grows, would be expected. Thus, we transformed the recall ratio of Experiment 1 from the portion of items correctly recalled in a category to the proportion of the items recalled of a certain semantic category in the total number of items recalled correctly. This transformation was based on an assumption that the portions of recalled items of animal and artifact in the total recalled items were not altered by the list length in the case of the mixed lists being used for learning. We compared our data from Experiment 1, with 32 items referring to animals or artifacts, to the data of Bonin et al. (Reference Bonin, Gelin, Laroche, Meot and Bugaiska2015), in which the animacy effect was found in five experiments with list lengths of 28 items and 56 items. Results of a multivariate analysis of variance (MANOVA) indicated that portions of either animate or inanimate items recalled in the total items recalled were not altered by the list length, F(2, 237) = 2.272, p > .05, 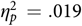 ${ { {\eta _p^2} \,}}. = .019$. This result indicated that the portions of both animate and inanimate items recalled in the total number of recalls were independent of the list length. Thus, we could infer the recall ratio of both animals and artifacts when the list length was 16 items as they were memorized in the themed list. This result allowed us to compare the data from Experiment 2, in which themed lists were used, to the data from Experiment 1, in which mixed lists were used, although the list lengths were not equal.
${ { {\eta _p^2} \,}}. = .019$. This result indicated that the portions of both animate and inanimate items recalled in the total number of recalls were independent of the list length. Thus, we could infer the recall ratio of both animals and artifacts when the list length was 16 items as they were memorized in the themed list. This result allowed us to compare the data from Experiment 2, in which themed lists were used, to the data from Experiment 1, in which mixed lists were used, although the list lengths were not equal.
The transformed data are presented in Table 3. The memory performances of NRP items of animals were not different between the two experiments (Experiment 1: M = 0.577, SD = 0.129; Experiment 2: M = 0.609, SD = 0.133), t(94) = −1.136, p > .05, d = −0.246, but a strong resurgence of the memory of NRP items of artifacts was observed in Experiment 2 (Experiment 1: M = 0.423, SD = 0.129; Experiment 2: M = 0.586, SD = 0.186), t(94) = −5.016, p < .001, d = −1.086 (see Figure 3a). The Bayes factor analysis revealed the same results, with anecdotal evidence to support the absence of memory recovery of animals, BF01 = 2.520 (see Figure 3c), but extremely strong evidence to support the memory of artifacts was recovered in Experiment 2, BF01 = 0.000 (see Figure 3d). No significant difference was observed between the two experiments of the memory of either animate or inanimate RP- items, t(94) = −0.238, p > .05, d = −0.052 for animals, t(94) = 1.291, p > .05, d = 0.280 for artifacts (see Figure 3b), and BF01 = 4.317 for animals, BF01 = 2.140 for artifacts (see Figure 3e and f).
Table 3. The descriptive statistic of the portion of correct recall transformed from Experiment 1
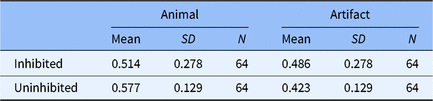
Fig. 3. Results from the comparison between experiment 1 and experiment 2 analyzed using: (a) null significance hypothesis testing for the memory recovery of NRP items; (b) null significance hypothesis testing for the memory recovery of RP- items (error bars denote ± 2 SE) and sequential analysis from Bayesian independent t tests for (c) the memory recovery of NRP items of animal; (d) the memory recovery of NRP items of artifact; (e) the memory recovery of RP- items of animal; (f) the memory recovery of RP- items of artifact.
Discussion
Consistent with our prediction, the results indicated the absence of animacy effect when animals and artifacts were divided into two memory sets. There was no difference in the memory of RP- items of either animal or artifact between the two experiments. Moreover, the memory of the NRP items of artifacts in Experiment 2 was much better than it was in Experiment 1. These results suggest the recovery of the memory of NRP items of artifacts when the memory set consists of items belonging to the same semantic category. Therefore, the absence of animacy effect is the consequence of the memory recovery of NRP items of artifacts rather than the decreasing memory of animals. The memory of NRP items of artifacts seems to be altered by the memory of animals. The potential mechanism may be that the inhibitory control of retrieval selects artifacts as the targets to suppress in order to facilitate the retrieval of animals. Therefore, by eliminating the inhibition that comes from the prepotent category (animal), the memory of NRP items of artifacts is recovered. With the resurgence of the recall of NRP items of artifacts, the animacy effect is eliminated. The animacy effect is more likely due to the suppression of the memory of other categories in the mixed memory set.
The results also illustrated the extent to which the animals and artifacts were influenced by the inhibition control in the case of only within-category inhibition. Consistent with our prediction, a strong inhibitory effect on artifacts was obtained, suggesting the memory of artifacts was impaired by the retrieval practice. Though the results indicated an absence of inhibitory effect on animals, the results of Bayes factor analysis showed an anecdotal evidence (BF01 = 1.208). It is not appropriate to confirm that animals were not influenced by inhibitory control based on such evidence. Altogether, the animacy effect may depend on the selective inhibition on the vulnerable categories during the retrieval process rather than the insensitivity to inhibitory control.
General discussion
In this study, we have examined the role of inhibitory control in animacy effect. A strong animacy effect and an inhibitory effect on animate items was obtained in Experiment 1. These results show that animacy effect is not due to the insensitivity of animate stimuli to the inhibitory control. Contrary to the prediction of the well-established theory of inhibitory control, we have found that the inhibitory control has no effect on the memory of inanimate items. The second experiment was conducted to test the role of inhibitory control when animals and artifacts are divided into separate memory sets, and the results showed a strong inhibitory effect on artifacts but the moderate evidence to support the absence of the animacy effect.
The results of two experiments indicate that the memory of both animals and artifacts is impaired by the retrieval practice, which represents the effect of inhibition in the memory process. Despite the absence of inhibitory effect on animate stimuli in the second experiment, evidence from the Bayes factor test was not sufficient to support the assumption that inhibitory control had failed to have an effect on animate items. The RIF effect observed in both experiments demonstrated that animacy effect is not due to the insensitivity of inhibition, but to the suppression of other semantic categories competing for retrieval. Although the role of inhibitory control does not exert an equal effect on the animals and artifacts, it seems to play a fundamental role in animacy effect in the condition of selective retrieval competition.
Animate stimuli are more likely to be the unintended prepotent targets during the retrieval process. The inhibitory control may be sustained without any intention of avoiding the unwanted memory if retrieving diversionary items becomes habitual (Anderson & Green, Reference Anderson and Green2001). Though animate stimuli were not the explicit target in our experiments, the inhibitory control process had a selective effect on the other category that ensured the animals would be retrieved for an adaptive purpose. Nairne (Reference Nairne2010) suggests that humans know the world by a “Stone-Age brain”. Animate stimuli are more important than other stimuli for the survival competition, therefore the “Stone-Age brain” always retrieves animals into awareness, even in the modern age. Previous studies have provided evidence that people tend to recall animate items even if they do not need to. For example, Bonin et al. (Reference Bonin, Gelin, Laroche, Meot and Bugaiska2015) found that animate stimuli are remembered better in the condition when the animacy dimension is not made explicit to the participants. A more extreme example is the study of VanArsdall et al. (Reference VanArsdall, Nairne, Pandeirada and Blunt2013), where they presented some pronounceable non-words (e.g., “frav”) associated with animate or inanimate properties to participants; the results indicated a great memory advantage for the non-words that are contaminated with animate properties. These studies suggest that animate stimuli tend to be retrieved more preferentially than inanimate stimuli during the selective retrieval process, even when there is no need to compete with other categories.
Since animate stimuli are retrieved preferentially, other stimuli become the target that need to be suppressed to reduce their competition for retrieval. In most cases, human memory contains multiple categories of items. When a category is more suitable for the current situation, the items in the category become the prepotent targets competing for retrieval with others. For example, Gelin et al. (Reference Gelin, Bugaiska, Meot and Bonin2017) have reported a significant animacy effect on recall in a survival scenario but absence in a non-survival situation. They argue that people pay more attention to the inanimate objects in the non-survival scenario. An alternative explanation of their results is that inanimate objects may be more suitable for the non-survival situation. Therefore, inanimate objects become the retrieval targets in the non-survival scenario, and the inhibitory control is turned to the unwanted animals during retrieval competition. In addition, a decline of memory of animals is observed in the non-survival scenario compared to the survival scenario in their study, which provides further evidence to this end.
The inhibitory control account is also able to explain the absence of animacy effect in cued recall tasks. Popp and Serra (Reference Popp and Serra2016) found that animacy enhances free recall but impairs cued recall. The inanimate items are remembered better than animate items in their three experiments when the final tests are conducted by using the cued recall tasks. The animacy seems to have a negative effect on memory in paired-associate scenarios. They argue that the attention and mental arousal should be the moderators between animacy and memory. Their results can also be explained in the framework of inhibitory control. The mechanism of inhibitory control in the process of paired-associate learning may differ from the mechanism in free recall. The cued recall is influenced by item memory for the probe and target words in addition to memory for the association itself (Madan, Caplan, Lau, & Fujiwara, Reference Madan, Caplan, Lau and Fujiwara2012). In the cued recall task, the memory of items is competing with the memory of the associations for retrieval. Though the associations are explicit to participants, the individual animate items are the prepotent memories for retrieval at the cost of other information that is selected to be suppressed during the retrieval competition. In other words, the memory for animate items per se is retrieved by inhibiting the memory of the associations between items.
In summary, the animacy effect is the fact that the animate stimuli are more likely to be retrieved compared to other stimuli. It seems that human memory system will preferentially retrieve animate stimuli at the cost of other stimuli. The role of inhibitory control in animacy effect is to suppress the retrieval of other categories. Due to the pressure of evolution, animate stimuli become the prepotent targets to be retrieved in order to fit the survival environment. Thus, inanimate objects are selected to be inhibited to ensure the retrieval of animals. This mechanism is formed in a long history of evolution, ensuring that humans can succeed in survival competitions. People do not need to pay extra attention to animate stimuli in the modern age, but the feature of preferential retrieval of animate information is preserved. The inhibitory control on inanimate objects may be an automatic process when they are presented along with animate stimuli. Although our inference cannot completely account for the mechanism of animacy effect, it is close to the proximate mechanism.
Author ORCIDs
Jun Tao, 0000-0001-6717-5524
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/prp.2019.20
Acknowledgements
The authors thank Dr Yanchao Bi and the anonymous reviewers for their constructive comments on previous versions of the manuscript; also, thanks to Dr Shichao Ma for advice on the statistical methods. The present study was supported by the Department of Education, Heilongjiang Province, China (GBC1317069).
Appendix
Materials and Norms in the Present Study
Materials and Norms in the Present Study








 Open access
Open access