



Introduction
T. spiralis infection is a common helminthic zoonotic disease with worldwide distribution (CDC 2017). It affects a wide range of mammals, birds, and reptiles. Man can be infected by ingesting T. spiralis larvae in raw or undercooked meat like pork and bears (Wang et al. Reference Wang, Liu, Sun, Song, Jiang, Zhang and Cui2017). Globally, T. spiralis has infected about 11 million people, and with recent outbreaks recorded in 55 different countries, it has gained attention as a public health problem (Pozio Reference Pozio2007; Luis Muñoz-Carrillo et al. Reference Luis Muñoz-Carrillo, Maldonado-Tapia, López- Luna, Jesús Muñoz- Escobedo, Armando Flores-De La Torre, Moreno-García and Bastidas2019).
Trichinellosis in humans can be divided into intestinal and muscular phases. Acute intestinal infection usually starts with non-specific manifestations like headache, fever, and gastroenteritis with diarrhea (Gottstein et al. Reference Gottstein, Pozio and Nockler2009). Chronic trichinellosis commonly occurs 3 to 4 weeks after ingesting larvae; it can cause persistent numbness, increased sweating, decreased muscle strength, and conjunctivitis. It can last up to 10 years post-infection in those who were not treated promptly during the acute stage of the infection (Gottstein et al. Reference Gottstein, Pozio and Nockler2009; Sun et al. Reference Sun, Wang, Liu, Jiang, Liu, Wen, Qi, Wang and Cui2015). Complications like respiratory failure, myocarditis, and encephalitis may occur (Yu & Qi Reference Yu and Qi2015).
T. spiralis infection causes pathological alterations in a variety of tissues, including the small intestinal epithelium and the skeletal muscles. Tissue damage from trichinellosis is caused by several mechanisms other than the direct harm caused by the parasite itself. The oxidative stress that comes along with trichinellosis is one of the primary causes of this damage, as shown by the increased production of numerous stress markers, including malondialdehyde (MDA) and superoxide dismutase (Bruschi et al, Reference Bruschi, Saviozzi, Piaggi, Malvaldi and Casini2003; Mido et al. Reference Mido, Fath, Farid, Nonaka, Oku and Horii2012).
Antioxidants may defend the host against oxidant-induced injury and the negative effects of molecules formed as a result of defense responses by the host (Abdallahi et al. Reference Abdallahi, Hanna, De Reggi and Gharib1999). Experimental trichinellosis has been shown to alter the activity of some antioxidants, such as superoxide dismutase, glutathione peroxidase, and glutathione-S-transferase (GST) in the muscular tissue (Derda et al. Reference Derda, Wandurska-Nowak and Hadas2004).
Traditional anti-Trichinella spiralis medications such as albendazole and mebendazole are effective against intestinal stages but less effective against encapsulated larvae. In addition to the growing resistance against these drugs, they have low water solubility, which restricts their absorption from the intestine and reduces their bioavailability (Khalikov Reference Khalikov2021). Accordingly, they are utilized in large dosages with negative side effects (de la Torre-Iglesias et al. Reference de la Torre-Iglesias, Garcia-Rodriguez, Torrado, Torrado, Torrado-Santiago and Bolas-Fernández2014). As a result, there is an urgent requirement for new and safer medications, particularly those that are effective against larval stages of Trichinella, which can result in serious types of long-term inflammatory myositis and myopathy.
ZnO is one of the five zinc compounds that the Food and Drug Administration considers safe for human use (Premanathan et al. Reference Premanathan, Karthikeyan, Jeyasubramanian and Manivannan2011). Zinc oxide nanoparticles (ZnO NPs) can enhance the action of numerous serum enzymes and improve the oxidant status (Ahmadi et al. Reference Ahmadi, Ebrahimnezjad, Ghiasi and Sis2014). These NPs have good biocompatibility with human cells and antimicrobial activity. They disrupt the bacterial metabolic processes and damage microbial cell walls, resulting in their death (Chitra & Annadurai Reference Chitra and Annadurai2013; Siddiqi et al. Reference Siddiqi, Rahman, Tajuddin and Husen2018).
Therefore, this study is conducted to evaluate the antihelminthic effect of ZnO NPs and their combination with albendazole on intestinal and muscular phases of trichinellosis and their effect on plasma oxidant/antioxidant status and vascular endothelial growth factor (VEGF) gene expression in muscle tissue.
Materials and methods
Experimental animals and T. spiralis infection
Laboratory-bred Swiss Albino male mice weighing 20–25 gm, free of pathogens, were used. Mice were obtained from the biological supply center of Theodor Bilharz Research Institute (TBRI) in Giza, Egypt. The study was conducted in the Medical Parasitology and Medical Biochemistry Department laboratories at Zagazig University, Egypt. T. spiralis larvae for mice infection were obtained from TBRI. Larvae were consistently maintained in the laboratory by being consecutively passed in mice. According to Dunn and Wright (Reference Dunn and Wright1985), T. spiralis-infected mouse muscles were cut and digested in a solution formed of 1% pepsin and 1% concentrated hydrochloric acid in warm tap water. After overnight incubation at 37°C, larval extraction was done by the sedimentation method, followed by washing many times with saline. A hemocytometer was used to count the number of larvae per ml. Each mouse was orally infected by 250–300 T. spiralis L1 larvae.
Ethics guidelines
The protocol of the current work was approved by the ZU-IACUC committee at Zagazig University (Approval number: ZU-IACUC/3/F/150/2022).
Experimental design
Mice were divided into five groups (ten mice each): Group 1, non-infected control; Group 2, infected control; Group 3, infected and treated with albendazole; Group 4, infected and treated with ZnO NPs; and Group 5, infected and treated with a combination of albendazole and ZnO NPs.
Control groups (1 and 2) were subdivided into two subgroups (A and B). Similarly, infected treatment groups were equally subdivided into two subgroups (A and B) according to the time of starting treatment. Treated groups (A) started treatment from the 2nd dpi, and treated groups (B) started treatment from the 35th dpi. To evaluate the effect of drugs on the enteric phase, mice of subgroups (A) were sacrificed at the 7th dpi. For the muscular phase, mice of subgroups (B) were sacrificed at the 45th dpi (Elmehy et al. Reference Elmehy, Ismail, Soliman, Amer, Elkaliny, El-Ebiary and Gamea2021).
Drugs
Albendazole treatment
Albendazole was administrated orally at a dose of 50 mg/ kg for 3 consecutive days. Alzental suspension (Albendazole 20 mg/ml) from EIPICO, Egypt, was used (Attia et al. Reference Attia, Mahmoud, Farrag, Makboul, Mohamed and Ibraheim2015).
ZnO NPs treatment
ZnO NPs were obtained from Nano Gate Company, Cairo, Egypt. NPs were characterized by transmission electron microscopy (TEM) equipped with high resolution (JEOL JEM-2100, JEOL Ltd., Tokyo, Japan) and by Zetasizer Nano ZS (Malvern Instrument Ltd., Malvern, UK). Mice were intraperitoneally injected with ZnO NPs dissolved in normal saline (0.9%) in a dosage of 5.6 mg/kg/day for 5 days (Bauomy Reference Bauomy2020).
Parasitological studies
Isolation and counting of adult worms
Animals were sacrificed, and the small intestines were removed. After washing the intestines with physiological saline, they were cut into 1-cm sections and maintained in physiological saline for 3 h at 37°C. The saline was collected, and intestinal sections were washed three times with physiological saline. The entire fluid was collected and centrifuged at 2000 rpm for 3 min. After discarding the supernatant, the sediment was reconstituted in 3–5 drops of physiological saline. The adult counts were detected using a dissecting microscope at a magnification of 20x (Wakelin & Lloyd Reference Wakelin and Lloyd1976).
Total larval counts in muscles
Muscle larval counts in whole corpses were detected according to the procedure outlined by Dunn and Wright (Reference Dunn and Wright1985). In brief, each mouse was dissected and digested in 1% pepsin-hydrochloride in 200 ml of distilled water. After mixture incubation at 37°C for 1 h with continuous stirring using an electric stirrer, larvae were collected by the sedimentation method and washed several times in distilled water. The count of larvae was determined by microscopy using a McMaster counting chamber.
Biochemical analysis
Tissue collection and preparations
Mice blood was taken into ethylenediaminetetraacetic acid and stored at 4°C. Muscle samples were washed with ice-cold saline, divided into small parts, weighed, and then homogenized with 50 mM phosphate buffer. This mixture was centrifuged, and the supernatant was stored at −80°C until usage (Belle et al. Reference Belle, Dalmolin, Fonini, Rubim and Rocha2004).
Assessment of oxidant/antioxidant status in plasma
GST and MDA levels were measured by Biodiagnostic kits (Giza, Egypt) according to the manufacturer`s recommendations. A colorimetric assay was done using the NanoDrop® ND-1000 Spectrophotometer (NanoDrop Technologies; Wilmington, Delaware, USA). Biochemical assays of GST and MDA activities were analyzed as described by Habig et al. (Reference Habig, Pabst and Jakoby1974) and Yagi (Reference Yagi and Yagi1982), respectively.
qRT-PCR for VEGF gene expression in infected muscle tissue (Schmittgen & Livak Reference Schmittgen and Livak2008)
Using Trizol (Invitrogen; Thermo Fisher Scientific, Inc.), total RNA was extracted from tissue homogenate. For testing the RNA quality, the A260/A280 ratio was done by the NanoDrop® ND-1000 Spectrophotometer (NanoDrop Technologies; Wilmington, Delaware, USA). The reverse transcription step and formation of complementary DNA (cDNA) were done by HiSenScriptTMRH [-] for the cDNA Synthesis Kit, INtRON Biotechnology, China. Evaluation of gene expression was done by qRT-PCR using 5 uL of the cDNA, 10 pmol/uL of each primer (1 uL each), and 10 uL of SYBR Green 2x Master Mix Green (QuantiTect SYBR Green PCR Kits, Qiagen). RT-PCR was achieved in an Mx3005P RT-PCR System (Agilent Stratagene, USA) following the manufacturer’s instructions. The PCR cycling conditions included an initial denaturation at 95°C for 12 minutes followed by 40 cycles of denaturation at 95°C for 20 seconds, annealing at 55°C for 30 seconds, and extension at 72°C for 30 seconds. The oligonucleotide-specific primers were formed by Sangon Biotech (Beijing, China). GAPDH was used to normalize our data, and the relative expression was calculated by the 2-ΔΔCt method (Schmittgen & Livak Reference Schmittgen and Livak2008). Table 1 shows the primer sequences of our research.
Table 1. VEGF, GAPDH primer sequences and conditions of amplification

b RTPrimerDB (RT-PCR Primer and Probe Database).
Histopathological study
Intestinal and muscle tissue specimens were initially fixed in 10% buffered formalin. After washing in distilled water, samples were dehydrated in ethanol, cleared in xylol, and then embedded in soft paraffin. 5 μm-thick sections were then stained with hematoxylin and eosin (H&E) (Alturkistani et al. Reference Alturkistani, Tashkandi and Mohammedsaleh2016).
Statistical analysis
The data were analyzed using SPSS version 25 (IBM, Armonk, NY, USA). Quantitative data is represented as mean ± SD. The ANOVA (F) test was used to compare three or more groups with quantitative values; pairwise comparisons between groups were performed using the Least Significant Difference method. For qualitative data, the Chi-square test (χ2) was used. Statistical significance was defined as P value < 0.05.
Results
Characterization of ZnO NPs
TEM image of the ZnO NPs revealed that the prepared NPs were in nanoscale with an average size of 30± 5nm and a spherical shape (Figure 1). The zeta potential of prepared NPs was +15.4 mV, indicating the stability of ZnO NPs suspension with a positive charge (Figure 2).

Figure 1. TEM picture of ZnO NPs.

Figure 2. Zeta potential of ZnO NPs.
Parasitological results
Adult worm counts in small intestine at 7th dpi
There was a high statistically significant difference between all infected groups in the mean counts of adult T. spiralis (p < 0.001). The highest level of reduction was detected in G5A, followed by G3A, and then by G4A (100, 99, and 57%, respectively). All treatment groups showed significant reductions in adult worm mean numbers in comparison to G2A (p < 0.001); see Table 2.
Table 2. Adult worm counts and larval counts

a, b, c, d insignificant difference (p > 0.05) between any two groups in the same column with the same superscript letter. Groups with different letters are statistically significant (p < 0.05)
F: ANOVA test; ** p < 0.001: highly significant; R%: reduction percentage
Total larval counts in muscle tissue at 45th dpi
Larval counts in all treated subgroups significantly decreased compared to G2B (p < 0.001). G5B presented the maximum reduction of encysted larvae (83%). However, G4B showed a 49% larval reduction, but G3B showed a 68% reduction. Additionally, there was a significant statistical difference among inoculated subgroups (p < 0.001); see Table 2.
Biochemical results
MDA and GST Levels in plasma
MDA level in plasma was significantly increased in G2 in comparison to all groups in both intestinal and muscular phases (p < 0.001); see Table 3.
Table 3. Oxidant/antioxidant markers levels in plasma during intestinal and muscular trichinellosis

a, b, c, d insignificant difference (p > 0.05) between any two groups in the same column with the same superscript letter. Groups with different letters are statistically significant (p < 0.05)
F: ANOVA test; ** p < 0.001: highly significant
In contrast to the plasma level of GST, G2 showed a significant decrease in GST plasma level compared to all other groups (p < 0.001). There was a highly significant difference between all groups; see Table 3.
VEGF gene expression in muscle tissue
VEGF gene expression level was measured in muscular tissue homogenate of the different studied subgroups using qRT-PCR at 45 dpi. The mean of VEGF gene expression in G2B was significantly increased in relation to all other groups (5.06 ±0.36, p < 0.001). G5B showed the greatest level of inhibition in VEGF gene expression (1.14±0.05); see Table 4.
Table 4. Relative expression level of VEGF gene detected in muscular tissue in the different study groups

a, b, c, d insignificant difference (p > 0.05) between any two groups in the same column with the same superscript letter
F: ANOVA test; **p < 0.001: highly significant difference; I%: increase percentage
Histopathological results
Small intestinal histopathology
G2A showed intestinal tissue with irregular short villi and an expanded fibrovascular core by many mononuclear and polymorphonuclear inflammatory cells. Adult worm sections and a decrease in goblet cells were also detected. However, G3A showed slightly short broad villi, few cellular inflammatory infiltrations, and a mild decrease in goblet cells. In comparison, G4A presented regenerating villi with an oedematous fibrovascular core with a moderate number of inflammatory cells. G5A showed maximum improvement in pathological changes. It showed mostly regular villi of normal length with mild inflammation and a normal goblet cell population (Figure 3).
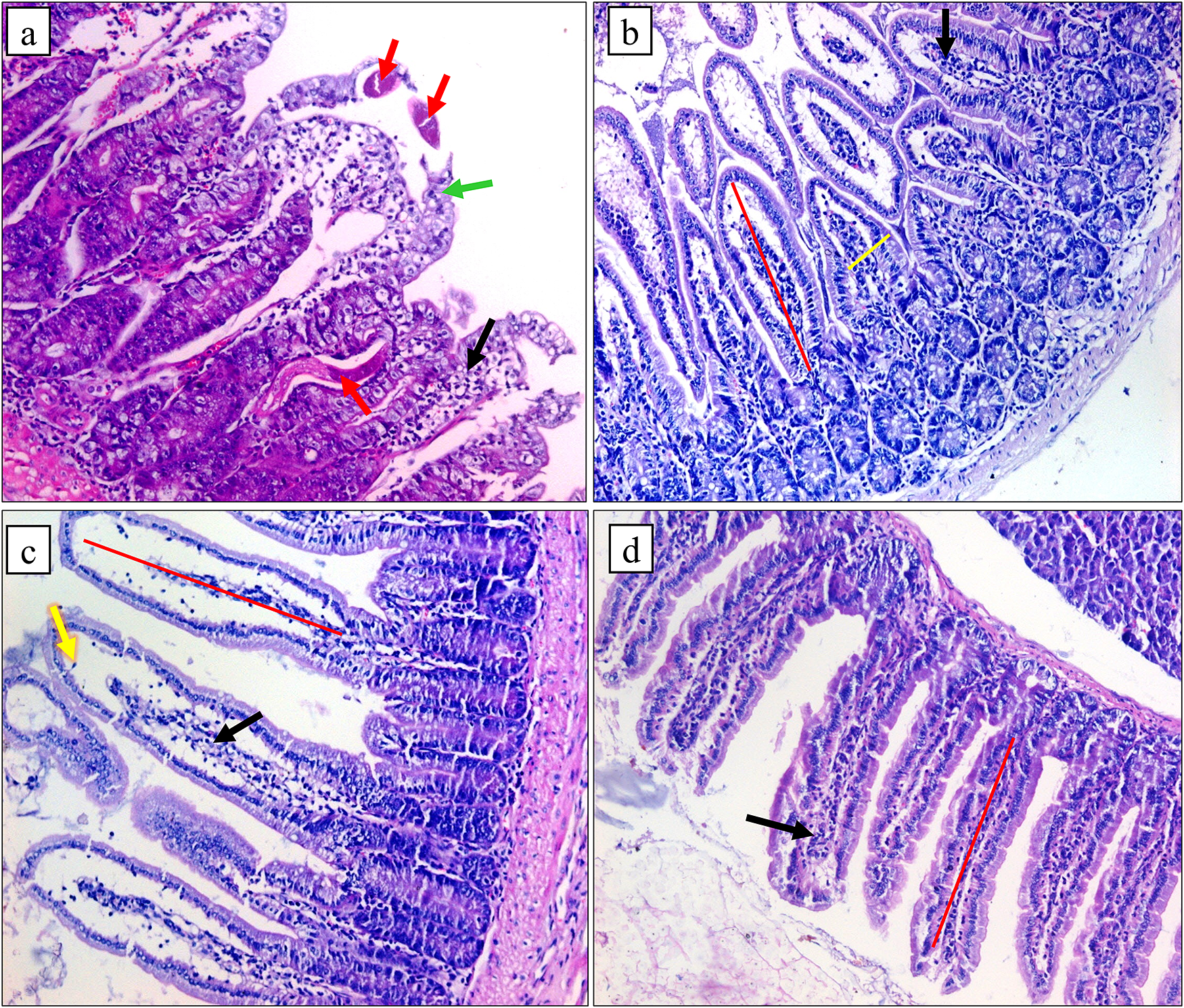
Figure 3. Histopathological pictures of intestinal sections (a) G2A showing adult T. spiralis worms (red arrows), irregular short broad villi (green arrow), and marked inflammation (black arrow), (b) G3A showing slightly short broad villi (red and yellow lines) and few cellular inflammatory infiltrations (black arrow), (c) G4A showing regenerating villi (red line) with oedematous fibrovascular core (yellow arrow) and a moderate number of inflammatory cells (black arrow), (d) G5A showing regular villi of normal length (red line) with mild inflammation (black arrow) besides normal goblet cell population (H&E x 200).
Skeletal muscle histopathology
Muscular tissue of G2B showed a large number of T. spiralis encysted larvae with diffuse muscle degeneration and dense infiltration by inflammatory cells. In contrast, G3B showed a decrease in the number of larvae with focal capsular and larval degeneration. Moderate infiltration by inflammatory cells was also detected. In G4B, there were some T. spiralis cysts with patchy muscle degeneration, infiltration by inflammatory cells, and focal larval degeneration. However, mice in G5B showed degenerated cysts with degenerated contents and few mononuclear inflammatory cells (Figure 4). Level of pathological changes is presented in Table 5.

Figure 4. Histopathological pictures of muscular sections (a) G2B showing a large number of T. spiralis encysted larvae with diffuse muscle degeneration (black arrows) and dense infiltration by inflammatory cells (red arrow), C: capsule, L: larva, (b) G3B showing a number of larvae with focal capsular and larva degeneration (black arrows) with inflammatory infiltration (red arrow), (c) G4B showing T. spiralis cysts with patchy muscle degeneration (black arrows), infiltration by inflammatory cells (red arrow), and fibrosis (green arrow), (d) G5B showing two small degenerated cysts with degenerated contents (yellow arrow), fibrotic capsule (black arrow), and few mononuclear inflammatory cells (red arrow) (H&E x 200).
Table 5. Extent of histopathological changes in intestinal and muscular phases in different groups

χ2: Chi-square test, * significant (p < 0.05)
Discussion
Trichinellosis is a highly prevalent and neglected tropical parasite in the world. It infects several species, including humans, and leads to major health issues (Robertson Reference Robertson2018). Benzimidazole compounds are frequently used to treat trichinellosis, but toxicity, side effects, low bioavailability, and resistance constrain the efficacy of those medicines. They also show limited reactivity to encapsulated Trichinella larvae. Thus, there is a growing need to identify and develop potent and safe anti-Trichinella agents (Fahmy et al. Reference Fahmy, Zalat and Rabei2020). Nanotechnology is a potential subject for creating innovative, efficient substances that range in size from 1 to 100 nm. Biocompatible NPs are one of the new methods investigated today for controlling parasitic infections (Elgadir et al. Reference Elgadir, Uddin, Ferdosh, Adam, Chowdhury and Sarker2015; Khan et al. Reference Khan, Singh, Ullah, Shoeb, Naqv and Abidi2015).
ZnO NPs are the third most widely produced nanometals worldwide (Piccinno et al. Reference Piccinno, Gottschalk, Seeger and Nowack2011). They were found to have no negative effects on human cells but to be hazardous to microbes (Reddy et al. Reference Reddy, Feris, Bell, Wingett, Hanley and Punnoose2007; Chitra & Annadurai Reference Chitra and Annadurai2013). Furthermore, ZnO NPs atoms provide more surface exposure, which causes them to display various physical, chemical, and extremely high levels of biological reactions (Wahab et al. Reference Wahab, Mishra, Yun, Kim and Shin2010). The current study is the first trial to evaluate the antihelminthic and antiangiogenic effects of ZnO NPs on T. spiralis infection.
In the current study, all treated mice showed a significant decrease in intestinal adults and encysted larvae in muscles compared to G2. Combination therapy produced complete eradication of intestinal Trichinella worms (100%) and the highest significant decrease in encysted larvae (83%). The albendazole effect was higher on T. spiralis adults than its effect on encapsulated larvae, with a percentage reduction of 99% and 68%, respectively. Similar results were obtained by Nada et al. (Reference Nada, Mohammad, Moad, El-Shafey, Al-Ghandour and Ibrahim2018) and Fahmy and Diab (Reference Fahmy and Diab2021), who detected more efficiency of albendazole against enteral stages of T. spiralis than parenteral ones. As noted, T. spiralis would become less vulnerable to albendazole when an infection becomes chronic, and its effectiveness against encapsulated muscle larvae is significantly diminished (Shalaby et al. Reference Shalaby, Moghazy, Shalaby and Nasr2010; Elmehy et al. Reference Elmehy, Ismail, Soliman, Amer, Elkaliny, El-Ebiary and Gamea2021).
In this work, treatment with ZnO NPs significantly affected both intestinal and muscular stages of trichinellosis (57%, and 49%, respectively). ZnO NPs have shown antihelminthic effects against various helminths, such as Toxocara vitulorum (Dorostkar et al. Reference Dorostkar, Ghalavand, Nazarizadeh, Tat and Hashemzadeh2017), Haemonchus contortus (Esmaeilnejad et al. Reference Esmaeilnejad, Samiei, Mirzaei and Farhang-Pajuh2018), Parascaris equorum (Morsy et al. Reference Morsy, Fahmy, Mohamed, Ali, El–Garhy and Shazly2019), and neuro-schistosomiasis mansoni (Bauomy Reference Bauomy2020). Also, it was effective against some protozoan parasites such as Leishmaniasis tropica (Meaad et al. Reference Meaad, Gharby and Al-Qadhi2017) and giardiasis (Brakat et al. Reference Brakat, El-Deen and Elhafiz2020).
Trichinellosis is associated with the production of high quantities of free radicals and reactive oxygen species (ROS) by the parasite and host as a defense mechanism in response to infection (Othman et al. Reference Othman, Abou Rayia, Ashour, Saied, Zineldeen and El-Ebiary2016). Antioxidants counteract the harmful effects of ROS, reducing damage and protecting body cells from oxidative harm (Banerjee et al. Reference Banerjee, Seth and Ahmed2001). In the current study, G2 showed a significant increase in oxidant marker (MDA) and a significant downregulation of antioxidant marker (GST) in plasma compared to G1 during both intestinal and muscular phases. Similarly, Abd El-Hamed et al. (Reference Abd El-Hamed, Abd-Rabou and Faramawy2022) found an increase in blood levels of the oxidant NOS2 and a decrease in α-GST in mice infected with trichinellosis. This was also in accordance with many other studies that detected increased oxidative markers and deceased antioxidants in tissues infected with T. spiralis (Kazemzadeh et al. Reference Kazemzadeh, Mohammad and Mohammad2014; Elgendy et al. Reference Elgendy, Othman, Hasby Saad, Soliman and Mwafy2020).
Oxidative stress was reduced in all treated mice, as presented by a decrease in plasma MDA levels and a rise in GST levels. Combination therapy presented the greatest improvement, followed by albendazole. In addition, ZnO NPs showed enhanced oxidative stress status during both intestinal and muscular stages of trichinellosis; they decreased MDA plasma levels and increased GST levels. Similarly, Nagajyothi et al. (Reference Nagajyothi, Cha, Yang, Sreekanth, Kim and Shin2015) reported that ZnO NPs showed antioxidant action by scavenging 2,2-diphenylpicrylhydrazyl (DPPH). In schistosomiasis, ZnO NPs significantly decrease MDA and increase glutathione levels in brain tissue, indicating their antioxidant effect (Bauomy Reference Bauomy2020). Treating murine trichinellosis with curcuma and pomelo NPs markedly improved all parameters in mice blood (Abd El-Hamed et al. Reference Abd El-Hamed, Abd-Rabou and Faramawy2022). Elmehy et al. (Reference Elmehy, Ismail, Soliman, Amer, Elkaliny, El-Ebiary and Gamea2021) also reported that treatment regimens appeared to boost GST action as one of the antioxidant participants in defense against T. spiralis.
Upon reaching muscular tissues, T. spiralis induces nurse cell formation and angiogenesis for parasite nourishment and protection from host immune responses. It increases the expression of VEGF, the chief angiogenic factor (Capó et al. Reference Capó, Despommier and Polvere1998; Kang et al. Reference Kang, Jo, Cho, Yu, Ock and Cha2011). The current study revealed a marked increase in VEGF gene expression in muscle samples of infected untreated mice in G2B compared to non-infected ones in G1B. Combination therapy showed the least gene expression, which was near that of G1B. This was in agreement with Khositharattanakool et al. (Reference Khositharattanakool, Morakote, Siriaunkgul and Uparanukraw2013), Othman et al. (Reference Othman, Abou Rayia, Ashour, Saied, Zineldeen and El-Ebiary2016), and Elgendy et al. (Reference Elgendy, Othman, Hasby Saad, Soliman and Mwafy2020), who proved upregulation of VEGF expression in muscles during trichinellosis and treatment with anti-Trichinella agents significantly reduced its expression related to positive control. ZnO NPs exhibited an antiangiogenic effect as they showed a significant reduction in VEGF gene expression compared to G2B. Similar findings were detected by Tada-Oikawa et al. (Reference Tada-Oikawa, Ichihara, Suzuki, Izuoka, Wu, Yamada and Ichihara2015) and Mohammad et al. (Reference Mohammad, Tabrizi, Ardalan, Yadamani and Safavi2019). They reported that ZnO NPs can significantly reduce the expression of VEGF-encoding genes. Gojova et al. (Reference Gojova, Guo, Kota, Rutledge, Kennedy and Barakat2007) also reported that ZnO NPs could stop angiogenesis by causing endothelial cell death.
Concerning the intestinal pathological study, combination therapy significantly reduced the severity of pathological alterations with restoring of normal villous architecture and normal goblet cells compared to G2A. ZnO NPs showed some anti-inflammatory effects; there were regenerating villi with moderate inflammation and a decline in goblet cells. As regards the histopathological changes in muscle tissues, the lowermost larval counts and least inflammation were detected in G5B. However, albendazole presented a moderate larval encapsulation in muscles with moderate inflammatory infiltration. In comparison, ZnO NPs showed a reasonable reduction in larval deposition with reduced inflammation. This was consistent with Shalaby et al. (Reference Shalaby, Moghazy, Shalaby and Nasr2010), Nassef et al. (Reference Nassef, Moharm, Atia, Brakat, Hussien and Shamseldeen2019), and Darwish et al. (Reference Darwish, MSA, Fahmy and FEA2022). It was stated that ZnO NPs have anti-inflammatory effects by suppression of iNOS, COX-2, IL-6, and TNF-α expression (Nagajyothi et al. Reference Nagajyothi, Cha, Yang, Sreekanth, Kim and Shin2015; Aboelmaatya et al. Reference Aboelmaatya, Omarab, Alya, Kotpa and Ali2022).
In conclusion, ZnO NPs decrease T. spiralis adult counts in the intestine and larval deposition in muscle. Additionally, they suppress VEGF gene expression in muscles and oxidative stress induced by trichinellosis. Also, their combination with albendazole enhances its effect on both the intestinal and muscular phases. This indicates that ZnO NPs have promising antihelminthic, antioxidant, and antiangiogenic effects against T. spiralis infection.
Financial support
Nil.
Competing interests
The authors declare no conflict of interests.









