This book is about scientific ideas and the evidence needed to exemplify and support the theory of evolution. It explores current biological diversity and asks the question how all the various life forms on our planet came about. Why do we have so many different species, and what processes cause biological change over geological time?
The Development of Evolution as a Science
An evolutionary narrative is often thought to begin with Charles Darwin, but historically evolutionary ideas have been with us for at least two millennia. Classical Greek philosophers such as Theophrastus (371 BC–287 BC) and Aristotle (384 BC–322 BC) were keen naturalists providing some of the first direct observations and empirical accounts of the natural world. And just as Theophrastus was studying plants (he was the first to systematically group plants) in lagoons nearby, Aristotle was contemplating the essential differences between plants and animals. Aristotle was interested in boundaries between species; not that he was presupposing speciation – for Aristotle believed in the ‘Ladder of Life’, a fixity of animal forms, moving from worms and simple creatures, through stages, to fish etcetera with Man at the top superseded only by the Gods. But Aristotle was prescient in that he saw nature as ‘changeable’ (in the manner rivers change the landscape over time) and ‘graded’ (as animals vary both from one another and from other animals), but species he believed were immutable and unchanging. Regarding the origins of life, he disagreed with both Empedocles (490 BC–430 BC) who had earlier suggested that life arose through chance assemblages in some early primordial soup and Anaximander (610 BC–546 BC) who speculated that all life arose in water. Charles Darwin himself thought Aristotle to be a proto evolutionist (not surprising as he was an acute observer of nature and keen to remove mysticism from the debate). But he was mistaken on this count due to an error made by a local town clerk who had mistranslated Aristotle’s ‘physics’. Aristotle was not supporting any species change but rebutting the argument put forward earlier by Empedocles. Darwin was not a classicist!
Later, as the classical texts of the Middle Ages gave way to the European Renaissance (fourteenth to seventeenth centuries) and then to the ‘Age of Enlightenment’ (eighteenth century), a profound shift in thinking was taking place. Encouraged by voyages of discovery around the world, wealthy individuals began to collect attractive and interesting specimens and display these within their ‘cabinets of curiosities’. Notably, the collection of Sir Hans Sloane became the basis of the collections now contained within the British Museum. Similarly, improvements in the technology of observation (telescopes and microscopes) together with developments in mass communication printing provided a further impetus for human intellectual voyages of discovery and with it the popularisation of science.
The process of collecting, cataloguing and displaying specimens eventually developed into a much more systematic endeavour. Collections of minerals and biological specimens were described and organised to uncover underlying organising principles. Explanations were also sought for the observations now being made. In truth, a scientific revolution was taking place where myth was to be replaced by theory, conjecture with evidence and simple curiosity with systematic investigation. Francis Bacon’s empirical approach led ultimately to ‘new ways of knowing’. Classical thinkers of the Middle Ages had been overtaken by what is referred to as the natural philosophers of the seventeenth and eighteenth centuries. Natural philosophy was a thoughtful and systematic study of the natural world. Subsequently the ‘scientist’ (a new term coined by Thomas Whewell in the mid-nineteenth century) would be associated with a practice involving hypothesis formation and rigorous testing of ideas. Charles Darwin (1809–1882) of course was an inspired scientist.
Darwin’s observations on biological complexity were systematic and his explanation of how this complexity arose took the form of a carefully reasoned argument. He used evidence to support his claims; evidence that could be checked and replicated by the wider scientific community. The earlier world views of Newton, Leibniz and Hobbes provided a rigid, almost clockwork view of the world, whereas in the mid-nineteenth century a more historical thinking prevailed. Examples of this new mind set include political thinkers such as Marx and Hegel who employed a dynamic and historical view of world events. Their thinking relates to a view of the world changing not the fixed view of their predecessors. Darwin’s half-cousin Francis Galton (1822–1911) had already explored increases in human population and its potential consequences while his grandfather, Erasmus Darwin (1731–1802), a prominent poet and biologist, alluded to a process of evolution and biological change in two of his long poems.
Contemporary with Charles Darwin, nineteenth century geologists such as Charles Lyell (1797–1875) and Adam Sedgewick (1785–1873) emphasised that the planet too was not a fixed entity but had undergone profound change ‘throughout the long expanse of history’. Limestone rock strata scattered throughout the British Isles demonstrated that these locations were once shallow seas with teeming marine life and not the Southern uplands and Yorkshire dales scenery that we see now.
The seventeenth-century image of an unmoving, static world was slowly being replaced by a more dynamic perspective. In the early nineteenth century, following the French revolution, there was a break with the more ‘classical’ approach. And proponents such as Lamarck and Saint-Hilaire challenged the (by now becoming outdated notion) of the ‘fixity of species’. This mind set affected Charles Darwin in his attempts to understand biological complexity. In 1859 Darwin published his On the Origin of Species by Means of Natural Selection together with his own description of biological change – ‘descent with modification’. Darwin also included a means by which these events could be explained, ‘natural selection’.
Charles Darwin was both a product of and contributor to this new way of thinking (or paradigm shift as Thomas Kuhn [Reference Kuhn1996] later called it).
As ‘natural philosophy’ gave way to ‘natural science’, a more rigorous, experimental approach or scientific method began to define scientific endeavour. Individuals such as Francis Bacon, 1561–1626 (philosopher, parliamentarian and scientist), Michael Faraday, 1791–1867 (the most eminent experimental chemist of his day) and William Whewell, 1794–1866 (President of the Geological Society) exemplified this approach. Whewell was a source of inspiration for Charles Darwin. Later that century biological science (the term ‘biology’ was coined in 1800 in an obscure German footnote) developed concepts such as the cell theory, principles of homeostatic control and impressive advances in animal and plant physiology through rigorous observation and experimentation. Biological evolution was slightly different, however. It did not at that time employ experimentation, but rather a systematic collection of evidence to answer questions together with an acutely reasoned argument. Following its synthesis with twentieth-century genetics, biological evolution rapidly became the cornerstone of biology; as Theodosius Dobzhansky famously says in his 1973 essay, ‘Nothing in biology makes sense except in the light of evolution’.
The history of evolution as an idea has had a long gestation, at times controversial, continuing in the twentieth century with development of evolutionary genetics. Genomics, a subject that did not exist before the twenty-first century, heralds a new chapter in our understanding.
The Years before Publication of Origin of Species
The year 1830, like many of those in the previous four decades, had been a turbulent one in French history. There had been revolution in Paris and the King was forced to abdicate. So when a friend called on the German poet Johann Wolfgang von Goethe in Weimar, he was prepared to agree that a great explosion had taken place in European affairs. But he was flabbergasted to discover that Goethe was referring not to French political upsets but to an acrimonious debate between two of the most noted comparative anatomists of the day, Georges Cuvier and Étienne Geoffroy Saint-Hilaire. For Goethe too was a considerable anatomist and appreciated the significance of the event.
The debate between the two former friends and current colleagues was not about evolution. The question, debated before a noisy audience in the premises of the Académie de Sciences in Paris, was about the correct way to interpret anatomical resemblances between different species of animals. To Cuvier, identity of structure meant identity of function; an animal, any animal, remained alive because it functioned like a well-coordinated machine. Every characteristic, internal and external, was created to serve its current way of life – no further explanation was required.
Geoffroy Saint-Hilaire agreed that functional anatomy was a worthy study, that anatomical features subserved a vital function. But to him functional anatomy was not a complete explanation. Quite apart from their function, the anatomical features suggested variation on an underlying plan. The proper task of ‘philosophical anatomy’ was to elucidate that plan – what, apart from their various ways of life, did all vertebrate animals have in common: could one reconstruct a basic vertebrate animal?
Over the years, Geoffroy Saint-Hilaire had attempted to implement this programme to the increasing irritation of Cuvier, but when Geoffroy Saint-Hilaire suggested that invertebrate animals, such as insects, lobsters and molluscs, also shared the same plans as vertebrates, open disagreement broke out.
Cuvier was a student of adaptation (that is the machine-like coordination of animal parts and the ‘fit’ of the whole animal to its environment), while Geoffroy was a student of homology (resemblances between species reflecting a common plan). Homology does not necessarily imply common ancestry, but it was due to the genius of Charles Darwin, through his Origin of Species, published in 1859, that both aspects of comparative biology were combined into a successful theory of evolution (Darwin, Reference Darwin1859).
So, What Is Evolution?
What do we mean by the term ‘evolution’? There are several different interpretations. Originally evolution implied some sort of unfolding, like the opening of a flower (Latin = evolutio: an unrolling), but latterly it has acquired a wider meaning, implying a general process of change. Darwin’s phrase ‘descent with modification’ accurately describes the process of biological change. This book is about biological (or organic) evolution – a system of theories put forward to explain both diversity and the relationships between different types of living thing.
If we wish to understand the theory of evolution, we need to consider the answer through a series of subordinate questions.
A theory is an established idea or organising principle used to explain a body of information. It covers a wide range of facts and forms and is said to possess both explanatory and predictive power. A theory is more than just mere speculation; a theory is a precise conceptual framework that supports the data. The theory of evolution by natural selection is a powerful explanatory tool. It makes predictions such as the existence of genetic variation (otherwise evolution could not happen) and patterns of speciation found in fossils (as seen in rock strata). It is supported by evidence from a range of sources, palaeontological, genetic, anatomical, behavioural and biogeographical; it even supports what Coyne (Reference Coyne2009, in his book Why Evolution Is True) refers to as retrodictions, facts and data that ‘make sense only in the light of the theory of evolution’.
In the construction of any theory there are two component parts:
1. the data to be explained (in philosophical terms we call this the explanandum) and
2. the theory or the explanation itself (the explanans).
So, what does evolution attempt to explain, what is its explanandum? Several answers have been offered to this question but there is a difference of emphasis among experts. Here are some possibilities.
The explanandum – evolution attempts to explain:
1. Why there are a staggering number of different types of living things alive on Earth today (some 30 million possible species)?
2. How it is possible to classify organisms in a hierarchical grouping, in Darwin’s phrase ‘in groups within groups’. Is there something real about biological classification? Does it suggest genuine relationships?
3. How the fossil record chronicles the biota – a sum of all life forms over time.
4. Why organisms appear to be particularly well adapted to their environment.
From these four questions above stem different schools of evolutionary research. And in order to answer the four questions above we can suggest,
The explanans
(In the same order as the questions were posed these are):
1. Those wishing to explain biodiversity and the ‘staggering number of different types of living things’ are likely to be interested in speciation; the division, in time, of one species into two or more and the mechanisms by which this occurs.
2. Taxonomists, interested in the classification and the hierarchical grouping of organisms, are concerned not only with constructing classifications but also with reconstructing the history of life (to which others including palaeontologists and molecular biologists also contribute).
3. Palaeontologists study fossils and explore life forms in different geological periods and can comment upon rates of evolution.
4. It is probable that most evolutionary biologists are preoccupied with the origin of adaptations – the reasons why adaptation is adequate rather than perfect and whether all the characteristics of organisms should be explained by natural selection.
To answer our question therefore (so, what is evolution?) we might say that evolution is a process of biological change – a theory that attempts to explain biodiversity together with an explanation in terms of differential reproductive success.
In addition to these lines of research there is a newly important branch of evolutionary theory, that of the evolution of development (or ‘Evo-Devo’ as it is known to its practitioners). For many reasons current evolutionary ideas do not fully explain how the development of individual organisms evolved. But in recent years there has been an explosion of knowledge in the role of the genome in animal development and the application of this knowledge to evolutionary problems.
It should be clear from what has been said so far that not only are there several sets of data that can be explained by evolutionary theory, but there are also several types of explanation. Together these represent the multifaceted discipline of evolutionary biology.
Change and Species Formation
In studying evolution, one is inevitably exploring biological change, the formation of new species together with the extinguishing of others.
But change and dynamism appear to be features of the world in which we live. Over its four billion or so year history the Earth has undergone profound change in terms of its geology, its atmosphere, the landscape, the climate and its constituent biota. Indeed, change in the abiotic (non-living) world often precedes or even dictates change in the biotic. Further proof, if needed, that all aspects of the natural world are interwoven
Perhaps a more cogent argument arises when scientists look beyond our own planet for signs of life. This new science of exobiology (also referred to as Astrobiology) needs to consider how extraterrestrial life might present itself. It presumably will need to secure an energy source and it will need to carry out various processes including coordinated activity and reproduction, but importantly (for the argument presented here) life will be seen to evolve. Evolution, or hereditable biological change over time, is now generally seen as one of the handful or so major characteristics of living things. Professor Gerald Joyce at the Salk Institute in the United States is an astrobiologist and an expert in the field of in vitro evolution (recreating the biomolecules of early life). Perhaps he has provided us with the best definition of life:
A self-sustaining chemical system capable of Darwinian evolution
One of Charles Darwin’s greatest achievements is to suggest a mechanism for the observed biological change over time – and that is natural selection. His theory of natural selection is both simple and elegant. Yet it is not reducible to the conventional rules of physics and chemistry. In this respect the biological sciences may be considered as inhabiting two epistemological ‘spaces’; on the one hand, the sciences of genetics, physiology, medicine and neuroscience (disciplines that are reducible to physical laws) and on the other, behaviour, community ecology and evolution which are not. Evolution it is argued belongs to this latter branch of whole organism biology where possible emergent properties arise and different research paradigms are needed.
Natural History and Classification
Organising our knowledge of the natural world and naming objects is a characteristic of human societies. Allied to this peculiarly human activity is the search for order and a desire to explain the world as it appears to us. The biological discipline dealing with the classification or grouping of organisms is known as taxonomy; this forms part of a more general speciality known as systematics (a study of the types and diversity of organisms). Confusingly, some biologists – mostly botanists – refer to a classification as a ‘taxonomy’.
Nomenclature (the naming of organisms) is a highly prescribed business. Before organisms can be classified, it is essential to have an agreed naming system. This applies not only to the naming of species but because classification of organisms is always expressed as a hierarchical structure (‘groups within groups’), there must also be rules about the naming of higher ranks. The whole system is policed by various International Commissions, most notably one for Zoology and one for Botany. Until the early 1960s methods for classifying organisms were ill-defined despite the fact that systematists claimed they were producing ‘evolutionary classifications’. Methods were largely intuitive. But then there arose not one, but two methods of classification, both claiming to be uniquely objective. They are known as phenetics (originally called ‘numerical taxonomy‘) and cladistics. Their practitioners often became bitter rivals, while both poured scorn on the easy-going and intuitive evolutionary taxonomists. The dust has now settled, and methods related to both phenetics and cladistics are in use for different taxonomic purposes.
Natural history as an academic enterprise has a long and distinguished history in the United Kingdom. The oldest biological society in the world, The Linnaean Society of London, was founded in 1788 to honour the botanist (Carl Linnaeus), his works and his legacy – his efforts in systematising the living world.
Elsewhere in Britain natural history became more organised with the standard works on identification produced. These included John Ray’s Catalogus Plantarum Angliae and Martin Lister’s Historiae Animalium Angliae, both published in 1678. It was in Plant Science or Botany that the discipline of natural history was first formalised. This is not surprising given the relevance of plants and plant products to the early study of medicine. The Society of Apothecaries based in London not only initiated the famous Physic garden at Chelsea but also promoted field trips into the local countryside. The earliest of these excursions was in May 1620 (the date of the voyage of the Mayflower to the New World). The Aurelians, as the lepidopterists (butterfly hunters) of the day like to call themselves, were another early specialist society.
In the mid-eighteenth century, natural history was more of a fashionable subject than a scientific one. It was perhaps the Victorians in the nineteenth century who forged natural philosophy to become the precursor of the more academic disciplines of Biology and Geology. Charles Darwin’s seminal work (Origin of Species) in 1859 interestingly provided a unifying theory for both the plant and animal sciences.
In 1866 a Chair in Zoology and Comparative Anatomy was created at Cambridge University, and the Education Act of 1870 brought a breakthrough in the teaching of Elementary Science. Indeed, there was such a shortage of teachers that the eminent zoologist Thomas Henry Huxley was asked by the government to set about providing a ‘crash course’ for teachers in botany and zoology.
There are many clubs, associations and learned societies that have contributed to our knowledge of the natural world. Both amateur and professional biologists are employed in the study of flora and fauna, local and national. It is upon this knowledge base, prepared by the natural historian, that the modern disciplines of taxonomy, ecology, ethology and (ultimately) evolutionary biology are founded.
An early example of a natural historian exploring evolutionary theory is that provided by Canon Henry Baker Tristram, born in 1822. ‘The great Gun of Durham’, as he was known, was an authority on birds in Durham, Northumberland and Palestine. As president of the British Association and Canon of Durham University’s College, Tristram (described as ‘a close observer and diligent collector’) was one of the first people to accept, in print, Darwin’s theory of evolution. This he did in an article in 1859 (less than one year after the publication of Origin of Species) in the ‘Ornithology of North Africa’:
Writing with a series of about 100 Larks of various species from the Sahara before me, I cannot help feeling convinced of the truth of the views set forth by Messrs. Darwin and Wallace in their communication to the Linnaean Society … it is hardly possible I should think to illustrate this theory better than by the Larks and Chats of North Africa.
Tristram then proceeds to discuss ‘gradual modifications of colouration and anatomical structure’ where ‘in the struggle for life … a very slight change for the better … would give the variety that possessed it a decided advantage over the typical or other forms of the species’ (Tristram, Reference Tristram1859: pp. 429–430). These views were also expressed in his Presidential address to the Tyneside Naturalists Field Club. This was a brave act coming from an Anglican churchman, but indicative of the growing acceptance of evolutionary theory.
Exploring the Development and Progress of Life on Earth
Reconstructing the history of life is usually regarded as the task of evolutionary biologists in general and palaeontologists, whose discipline takes in aspects of both biology and geology. Essentially, palaeontologists collect and prepare (that is clean up) fossils and then try to make valid statements about the anatomy, ecology and even behaviour of the organisms their specimens represent. Most palaeontologists are taxonomists and attempt to say something about the historical significance of their fossils by including them in a classification that also embraces living species.
A further category of evolutionary biology is that of the ‘adaptationists’ (there does not seem to be a suitable collective noun). Many are particularly interested in the evolution of behaviour (including human behaviour) and term themselves ‘behavioural ecologists’ or ‘sociobiologists’. Their principal preoccupation is with testing or applying Darwin’s theory of natural selection to the anatomy, behaviour and ecology of animals.
One thing Darwin could not do was provide a valid account of heredity – the mechanisms by which the characteristics of one generation are passed on to succeeding generations. No one could blame him for that as the work of Gregor Mendel (and hence the beginning of modern genetics) was only ‘rediscovered’ in the year 1900. At first a number of scientists believed that Mendel’s conclusions refuted Darwin’s theory of natural selection. The two theories were happily reconciled in the late 1930s and early 1940s in the so-called ‘Synthetic Theory’ of evolution. This new synthesis (the Synthetic Theory or Modern Synthesis) proposed that variation was brought about by random events and that populations evolve by means of changes in gene frequency (e.g. those brought about by natural selection). The Synthetic Theory is sometimes called ‘Neo-Darwinism’, the revival of an older term with a somewhat different meaning.
Evolutionary change can occur both above and below the level of species. Genetic change within a population, or below the species level is referred to as microevolution (that is changes in gene frequencies, mutation etc.). It is possible to demonstrate microevolution. Macroevolution, on the other hand, is evolution above the species level, including speciation. Its phenotypic changes affect the lineage of organisms and the ultimate appearance of higher groups (for example, the evolution of insects and the appearance of land plants). Macroevolution takes place over a much larger time scale and its progress is inferred using various lines of evidence, fossil appearance, radiometric dating, chemical analysis and degrees of relatedness.
By the mid-1960s it became possible to study evolution at the molecular level. In studying proteins, it became apparent that there was a greater diversity of molecular form within populations than previously imagined. Techniques such as gel electrophoresis confirmed the amino acid sequences of these molecules, while rates of change led to the suggestion of the possibility of ‘molecular clocks’. Motoo Kimura (1924–1994), a Japanese population biologist, hoped to combine the discipline of population genetics with the newly emerging molecular data. What emerged was a realisation that the observed variation within groups was too large to be explained simply by natural selection. He therefore proposed an alternative hypothesis, that of the Neutral Theory of Evolution. In this he postulated that molecular evolution was driven not necessarily by Darwinian natural selection but by random, non-adaptive changes within the genome.
Results of molecular studies have proved to be increasingly important in understanding the evolution of life on Earth, while the neutralist–selectionist debate has proved to be a useful focus for studies of molecular evolution.
To summarise, therefore, the Earth is a rationally ordered physical and biological system in which changes occur.
In the mid-seventeenth century James Ussher, the archbishop of Armagh, stated that the Earth was created the night before Sunday 23 October in the year 4004 BC! He did this by carefully measuring biblical genealogies. By 1800, however, geologists had demonstrated that the Earth must be older (for instance by calculating the length of time it takes for an object with the mass of the Earth to cool down). And Darwin, like his mentor the geologist Charles Lyell, believed in the Principle of Uniformitarianism (an agreement that processes we see in the present day also occurred in pretty much the same way as they did in the past); both Darwin and Lyell believed in a continuous, gradual geological change. The continuity of geological events on Earth is mirrored by Darwin’s thoughts on organic evolution – a classic expression of this Principle of Continuity.
Famously, in 1831 her Majesty’s ship ‘Beagle’ sailed from Devonport with the young naturalist Charles Darwin on board. And, as we now know, studies on the habits of the cuckoo, extinct quadrupeds, distribution of land shells and birds of the Galapagos Archipelago all contributed to his landmark text Origin of Species some 30 years later.
The Galapagos Islands and Darwin’s Finches: A Case Study
The Beagle’s orders were to survey and map the coastline of southern South America, then, following the Galapagos visit, to sail west via Tahiti, New Zealand and Australia, making astronomical and other observations. Darwin’s brief was, as guest naturalist, to study the geology and natural history. He landed home at Falmouth on 2 October 1836, nearly five years after the Beagle’s departure. Darwin recorded that ‘in July (1837) I opened my first notebook for facts in relation to the Origin of Species, about which I had long reflected, and never ceased working on for the next twenty years’. His great work on evolution, On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life, was not published until 1859.
The Galapagos Islands, over the many years since Darwin’s visit, have acquired an almost mythical status in accounts of the development of his theory. Some popular myths have Darwin’s conversion to ‘transmutation’ (i.e. evolutionary change) occurring suddenly during his five-week stay on the Galapagos, but there is no evidence of this other than an ambiguous note written as he prepared a catalogue of his bird specimens from previous ornithological notes, nine months after leaving the Galapagos. He was referring to the mockingbirds (Mimus parvulus) collected from four of the islands: the specimens from Chatham and Albemarle he says appear to be the same, but the other two are different. On each island each kind is exclusively found; habits of all are indistinguishable.
When I see these islands in sight of each other, and possessed of but a scanty stock of animals, tenanted by these birds, but slightly differing in structure and filling the same place in Nature, I must suspect that they are only varieties. If there is the slightest foundation for these remarks the zoology of Archipelagos – will be well worth examining, for such facts would undermine the stability of Species.
Darwin had also been told by the English vice governor of the Galapagos that the giant tortoises (Chelonoidis nigra) differed consistently from island to island but took little notice and did not collect museum specimens of the tortoises while there. The only tortoises collected by anyone (except for two babies kept as pets) were eaten by the Beagle crew and the skulls thrown overboard! Indeed, because of their size, hardiness and longevity, the tortoise population on the islands would be decimated by pirates and whalers who embarked onto the islands for shelter and provisions. It is reckoned that more than 100 000 of these lumbering reptiles (the megafauna of the Galapagos) were removed by seafarers.
The Galapagos archipelago comprises 16 volcanic islands of differing ages with varying landscapes (Figure 1.1). The younger islands like Fernandina in the West are inhospitable with harsh, arid landscapes of volcanic ash and lava flows and little vegetation. The older islands like Santa Cruz to the East are clothed in vegetation and are the centre of the Galapagos’ famed biodiversity. The oldest islands like Espanola, around 4 million years old, are sinking into the ocean with erosion reducing the landscape to a flattened coastal remnant. The significance of this is that the diversity in animal and plant life is mirrored by the diversity in habitat. And since emerging from the Pacific Ocean ‘hot spot’ around four and a half million years ago, organisms have populated these islands (arriving from the South American coast) providing a unique insight into adaptive radiation and the evolutionary process.
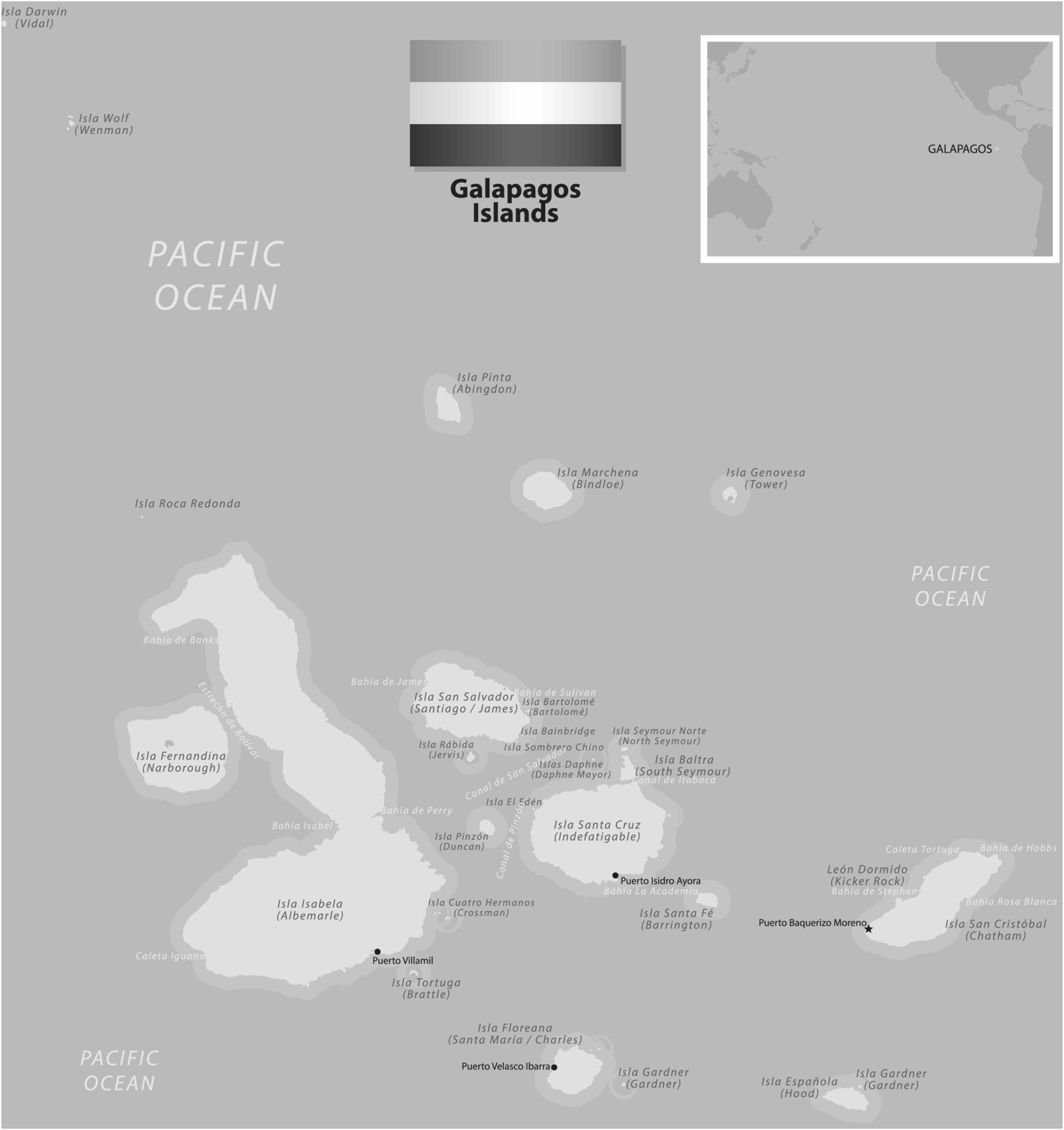
Figure 1.1 Sketch map of the Galapagos group of islands indicating the main sites referred to in the text.
Radioactive dating has shown that the oldest islands are about 5 million years old, the youngest about half a million. The importance of the isolation of the Galapagos from the South American mainland to the study of Darwin’s finches is that it seems there was never any land connection between the two areas. If a common ancestral species of all Darwin’s finches arrived from South America, it must have done so by chance – on floating mats of vegetation (which still detach themselves from coastal Ecuador today), by being blown off course or by other infrequent means. Thus, the founding population of the ancestral Galapagos finches presumably consisted of a small number of individual birds, with no doubt many casualties en route.
The environment of the Galapagos at first appears to the observer to be very inhospitable. Darwin’s first impression of ‘Chatham’ was not favourable:
Nothing could be less inviting than the first appearance. A broken field of black Basaltic lava is everywhere covered by a stunted brushwood which shows little signs of life. The dry and parched surface, having been heated by the noonday Sun, gave the air a close and sultry feeling, like that from a stove: we fancied even the bushes smelt unpleasantly.
Similarly, David Lack (1910–1973) speaks of ‘miles of dreary greyish brown thorn bush, in most parts dense, but sparser where there had been a more recent lava flow, and the ground still resembled a slag heap’. And yet the Galapagos have an enormously rich fauna, notably of birds, with many endemic species (unique to the island), and an equally rich marine life. A clue to the reason is the unique presence on the islands of a bird, whose group is more usually associated with Antarctica, the Galapagos penguin (Spheniscus mendiculus), the only equatorial penguin species. For islands on the equator the climate is often surprisingly cool and the sea conspicuously so. The explanation is the Humboldt Current which brings plankton-rich cold sea water up from the coast of Peru. The Galapagos climate is markedly seasonal. From roughly July to December, the Humboldt Current dominates, lowland air temperatures are cool and the rainfall slight. During this season, however, an inversion layer is created, and those islands with considerable highlands have those highland regions continuously wet, so they are covered in a rich green layer of plants. From January to June there is a warm, wet season in the lowlands with mostly clear skies but occasional heavy showers.
Before moving on to talk in detail about those finches, there is one more important point to be made. The birds are not quite unique to the Galapagos Islands. About 300 miles (500 km) southwest of Costa Rica in Central America, and nearly twice that distance northeast of the Galapagos, there is a small (47 km2) island called Cocos (situated on the Cocos plate). Like the Galapagos, it is a volcanic island, but in other respects it is very different. There is no seasonality: the whole island experiences heavy rainfall throughout the year and consequently is covered by dense rain forest. There are only four resident land birds on Cocos Island. One of those is a ‘Darwin’s finch’!
The vice governor’s comments about each island having its own type of tortoise extends also to other animals such as lava lizards, birds and various land snails. Each of these animals appears to have distinct forms unique to the different islands. Thus, one can postulate that the ancestral mainland ‘castaways’ that first arrived settled the various islands and therein adapted to their own unique environmental circumstances. So, tortoises on islands with lush vegetation developed high domed shells to move more easily (tracks made by tortoises in this dense vegetation are easily seen from the air), whereas those tortoises living where vegetation is poor acquired long necks and peaks at the front of their shells to allow them to reach up to the dominant plant (in this case the prickly pear cactus). In a similar vein, land snails in arid areas developed a small mouth (to avoid excessive water loss) and long, conical shells, while those snails in wet areas acquired wide mouths and globular shells. All of this points to a phenotypic plasticity and a rapid radiation of within-species forms adapted to its own environment and providing the potential for future speciation. But perhaps the most iconic example of island radiation and brisk speciation is the finches.
Current reckoning is that there are 14 species of finches on the Galapagos. Darwin landed on four of the islands during his five-week stay (although he saw many more as the Beagle criss-crossed the archipelago). He collected specimens from the four islands but attached so little importance to inter-island variation that he mixed up the specimens from the first two islands (Chatham and Charles) that he visited. Darwin did not even recognise that all the species formed a closely related group until the Beagle specimens had been studied by the Zoological Society’s ornithologist John Gould. Gould explained to Darwin that all the Galapagos finches in the Beagle collection formed a group of species more closely related to each other than to any other birds. Darwin’s conversion to transmutation seems to have occurred soon after this explanatory meeting with Gould in mid-March 1837. Nevertheless, there is no account of the finches in the Origin of Species of 1859, although Darwin does say of them in the second edition of his account of the voyage (1845), ‘One might really fancy that from an original paucity of birds in this archipelago, one species has been taken and modified for different ends’.
The Galapagos finches are small and mostly dull-coloured birds, little more than a sparrow size. The most striking feature of the whole group is the variation in the size and shape of the beak. The large ground finch Geospiza magnirostris has a massive parrot-like beak and a large heavily muscled head to support it. At the opposite extreme the little warbler finch, Certhidae olivacera, has a slender probing beak like that of a warbler! (Figure 1.2). Amazingly it took until the 1940s for general agreement that these beak shapes were adaptive, despite numerous early studies. This agreement resulted from the publication of an important book by David Lack, which popularised the phrase Darwin’s Finches as its title (Lack, Reference Lack1947).

The Finches
In the classification of organisms, the species is regarded as the fundamental taxonomic unit, and definitions of species (the ‘species concept’) have been a matter of vigorous debate since before Darwin’s time. Thus, any evidence bearing on the origin of a new species (or better still a whole series of new species) would be cogent evidence for evolution. The species concept is discussed in Chapter 6, but in general terms species are groups of freely interbreeding individuals separated from other groups by failure to interbreed. In a classification species are gathered together into genera (singular genus), but genera are to some extent arbitrary and subject to personal taste; there is little discussion, if any, about ‘the genus concept’.
It is thought that there are 14 species of Darwin’s finches on the Galapagos Islands, plus the one on Cocos Island. Not surprisingly large islands tend to have more resident species than smaller ones. Opinions vary as to the number of genera that should be used to group the finch species, but six genera are a generally acceptable number. They are as follows:
1. Geospiza: the ground finches
o Large ground finch – Geospiza magnirostres
o Medium ground finch – G. fortis
o Small ground finch – G. fuliginosa
o Sharp-beaked finch – G. difficilis
o Large cactus finch – G. conirostris
o Small cactus finch – G. scandeus
There is strong evidence, and general agreement, that the ground finches form a natural group. All are related to one another more closely than to any other species of Darwin’s finch. Technically the genus Geospiza is a monophyletic group, that is, the group consists of ancestral species and all their descendants.
2. Camarhynchus: the tree finches
o Large tree finch – Camarhynchus psittacula
o Medium tree finch – Camarhynchus pauper
o Small tree finch – Camarhynchus parvulus
The tree finches, as their name suggests, live much more in trees than do the ground finches. The tree finches are found more in the highlands and feed largely on insects.
3. Cactospiza: the woodpecker finches
o Woodpecker finch – Cactospiza pallida
o Mangrove finch – Cactospiza heliobates
The woodpecker finch is famous for not only using tools, but also for modifying them to its purpose. The behaviour was discovered in 1919 and has been observed frequently since. The bird uses a cactus spine, or a twig, often broken off by the bird itself, as a probe held lengthwise in the beak to winkle out grubs, etc. from cracks in bark. It also climbs up and down vertically like a woodpecker. The mangrove finch has also been observed in tool use. The former is a mostly highland species, the latter is found only in the mangrove swamps of Isabela.
4. Platyspiza crassirostris: the vegetarian finch
The only species in its genus; it lives in trees mostly in the highlands. Feeds on fruit, leaves and buds.
5. Certhidea: the warbler finches
o The green warbler-finch – Certhidea olivacea
o The grey warbler-finch – Certhidea fasca
The warbler finches feed mostly on insects, even in flight. Certhidea olivacea is found in the central, high islands (Santa Cruz and Santiago). C. fusca is known to occur in four lower more peripheral islands. Darwin did not accept that the warbler finch was any close relation of the others until convinced by Gould.
6. Pinaroloxias inornata: the Cocos finch
The single species from Cocos Island is small (13 g) with a slender, slightly curved beak like that of the warbler finch.
Despite uniformity in appearance, a study has shown that individual birds are specialist feeders with a large variety of ways of life in the rain forest. Specialities include searching for insects in leaves and in branches, looking for crickets and grasshoppers among dead leaves and collecting nectar.
Classification and the Galapagos Finches
Before any scientist can study objects, or phenomena, they must organise them some way. Ever since the days of the ancient Greeks, the most useful way of doing this has been to produce a hierarchical classification. Hierarchies come in two principal sorts. Both share the feature that they are defined by a series of ranks. The first is an exclusive hierarchy; an example here might be military rank. The second an inclusive hierarchy such as a taxonomic rank.
The term ‘rank’ perhaps needs explanation; in the words of the evolutionary biologist Ernst Mayr (1904–2005): ‘Military ranks from private, corporal, sergeant, lieutenant, captain up to general are a typical example of exclusive hierarchy. A lower rank is not a subdivision of a higher rank; thus lieutenants are not a subdivision of captains’.
In contrast though, a biological classification is an example of inclusive hierarchy. With Darwin’s finches, a single genus usually (but not always) includes several species. Then genera are grouped together in families and so on, at an ever more inclusive series of ranks (a set of nested groups). Biological classifications are usually also irregular – as an example, some genera have many species, others only one. By convention the hierarchy is also divergent; no species can belong to more than one genus.
In his book Darwin’s Finches (Reference Lack1947), David Lack not only presents a written classification of the birds (with discussion of the priority of the whole grouping within higher ranks) but also draws up a diagram looking like a family tree, with individual species at the end of each branch. He describes this diagram as ‘an evolutionary tree’, thus drawing an important theoretical conclusion, which is by producing a (correct) classification one is producing not just a diagram of that classification but also of the pattern of descent: an inclusive irregular classification is best explained as a phylogeny. Lack drew on previous work in drawing up his diagram; his classification was principally based on not only comparison of the appearance of the birds, but also of their ecology and behaviour such as song. If, however, one could use some completely different method and different data to classify Darwin’s finches and the new method produced the same result, then one could feel that the classification was ‘correct’. It would also show that the classification was in some way real and not just a convenient grouping of data. A ‘real’ classification based on natural groupings (birds, fish, insects, etc.) has been referred to as a natural classification, whereas a classification of convenience (all the waterfowl, all the yellow flowers) has been called an artificial classification.
Since Lack’s time, there have been numerous studies of Darwin’s finches, notably those of Robert Bowman from the 1960s to the 1980s including studies of beak function and song, which are outstanding not only as a series of works on the finches, but also as a thorough study of the ecology and evolution of animals. We can also note the research of husband and wife team Peter and Rosemary Grant (with a succession of colleagues, assistants and research students) from the 1970s to the present day.
In recent years the finches have been reclassified using techniques derived from biochemistry and molecular biology. In the 1970s and 1980s attempts were made at reclassification using the electrophoresis of proteins. But this method in general was not able to distinguish between individual species in such a closely related group of birds. With the 1990s came the use of DNA sequencing. Deoxyribonucleic acid (DNA) is the genetic material in all animals, present in the nucleus of all cells as the famous double helix, but also present as a single strand in the numerous mitochondria, the tiny power-houses of the cell, scattered through the cell cytoplasm. The genetic code itself consists of four bases (A: adenine, C: cytosine, G: guanine, T: thymine), an alphabet of four letters, anchored along the DNA molecule. But if two species of birds (or other organisms) diverge from one another over evolutionary time, point mutations can occur so that in one or both cases, one base is substituted for another at any site. The longer that two bird species have had a separate history, the more mutations are likely to occur, so that mutation number becomes a measure of elapsed time.
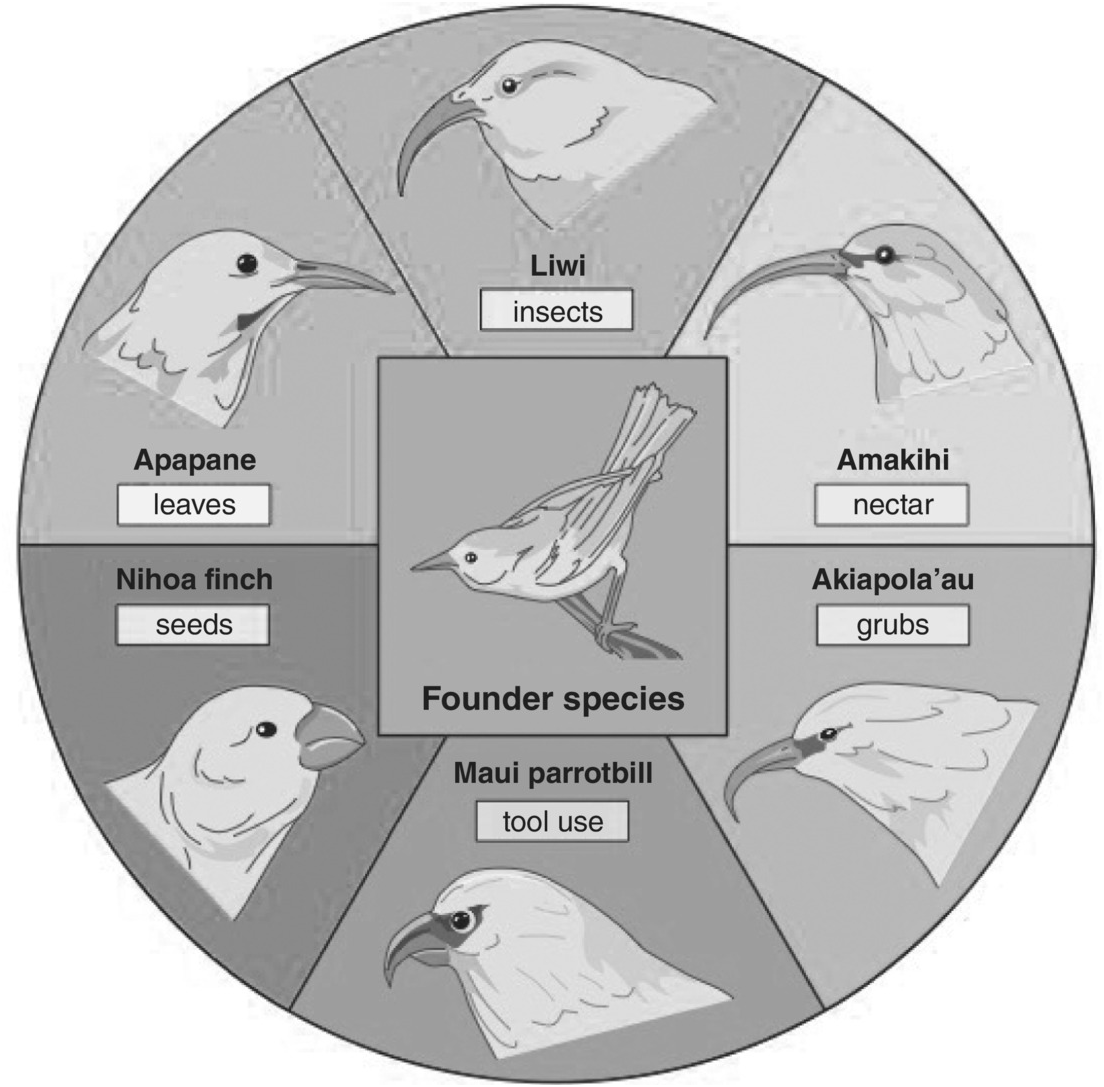
Both Galapagos finches and Hawaiian honeycreepers have been used to study adaptive radiation. Honeycreepers (see Figure 1.3) such as Apane (Himatione sanguinea), L’iwi (Vestiaria coccinea), Amakihi (Hemignathus virens), Akiapola (H. wilsoni) and the Nihoa finch (Telespiza ultima) also show the (relatively rapid) beak radiation of endemic island birds from a common ancestor.
Figure 1.3 The adaptive radiation of beak forms in Hawaiian Honeycreepers.
The hope is, that in comparing the DNA base sequence of one species with another, a ‘molecular clock’ will prevail, i.e. that the number of base differences in any sequence from two birds will be directly related to the time since the two diverged. This will not be true if the length of DNA has some vital function but might work for lengths of DNA of no known function.
Once again, the results with DNA sequencing of Darwin’s finches were short of convincing, but it became clear that the six ground finches were a natural closely knit group and that the warbler finches, comprising two species, had separated from all the others before the others had separated.
In these studies, pursued by members of the Grants’ ‘school’, both nuclear and mitochondrial DNA were used, but in the 1990s a new molecular technique came into use. In the nuclear genome there are frequently considerable lengths of repeated short (2 or 3) base motifs (e.g. …CACACACA…) of no known function. This is called ‘microsatellite DNA’. Mutation consists of the addition or deletion of individual motifs (a single CA, for instance). So what characterises the microsatellite DNA of a bird is not one or more point mutations of single bases, but microsatellite length. Comparison of one microsatellite length from each bird would be of little help, but large numbers of these microsatellites are available from any individual and the whole set characterises that individual (a similar technique is used in so-called genetic fingerprinting for forensic purposes).
In 1999, Petren, Grant and Grant were able to publish a phylogenetic tree of all species of Darwin’s finches, including the Cocos finch (Petren et al., Reference Petren, Grant and Grant1999). In many ways the satellite tree corroborates Lack’s tree and the ordering into genera as listed above. But there are important differences of pattern and interpretation, to some extent foreshadowed in the results of DNA sequencing. One is the separation of the vegetarian finch, which branched off the main stock before the ground finches and the tree finches (including the woodpecker finches) separated from one another. A second inference is that the Cocos finch is not the first to diverge from the Galapagos finches – one of the warbler finches has that honour. Therefore, the ancestors of the Cocos finch almost certainly colonised Cocos Island from the Galapagos and not from the South American mainland. And lastly, and perhaps most importantly, the warbler finches themselves are only distantly related to one another. C. olivacea was the first to diverge from the whole stock, then the Cocos finch, and then the other warbler finch, C. fusca. All this happened before all the remaining species became distinct. This leads to a most important conclusion. The resemblances between the two warbler finch species are not indicators of closeness of relationship and must therefore be primitive for all Darwin’s finches. They therefore probably give us a good idea of the appearance of the first-ever Darwin’s finch and are a guide to our search for the bird species most closely related on the South American mainland. Lack did not know that there were two species of warbler finch (although he did know that ‘it’ was divisible into several possible subspecies). He correctly suggested early divergence from the main stock but thought that the ancestor of the whole group would be like one of the ground finches, with black male plumage and a crushing beak.
In 2014 Skinner et al. explored epigenetic inheritance in five species of Darwin’s finch (see Figure 1.4). Epigenetic inheritance (more accurately transgenerational epigenetic inheritance) reflects the concept of heritable changes in which the physical structure of the DNA remains unchanged. The accepted model of course is that genetic mutation (established over time) generates the heritable phenotypic variation upon which natural selection acts. However, using erythrocyte DNA, this study explored the possibility that epigenetic changes can accumulate producing the ‘clay’ upon which natural selection may introduce change.
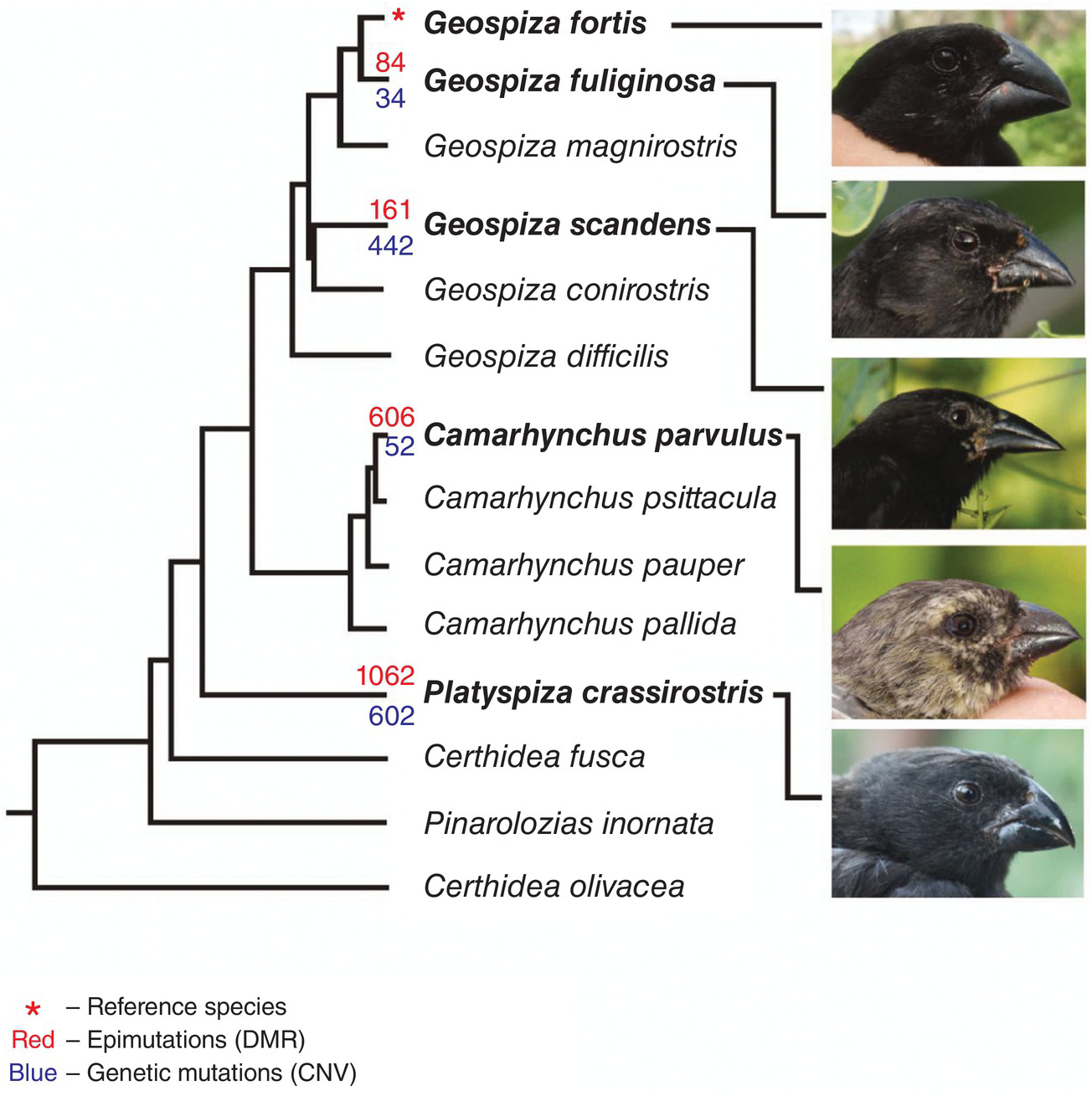
Figure 1.4 Numbers of genetic and epigenetic mutations in relation to the phylogeny of the Galapagos finches.
Blood samples were taken and analysed from five species of finch collected from Santa Cruz island in 2009. Genetic mutations were identified through use of gene duplication and deletion, while epigenetic variation was explored through differential DNA methylation analysis.
The phylogenetic tree in Figure 1.4 follows Lack’s physical traits reinforced by Petren, Grant and Grant’s microsatellite data. Epigenetic mutations (epimutations) are shown numerically (in red) along with copy number variation (genetic mutations) in blue. There were generally more epigenetic mutations than genetic ones indicating that epimutations may be a major component in evolutionary change. There was also a significant correlation between the number of epimutations and phylogenetic distance – perhaps suggesting that epigenetic change accumulates over long periods of evolutionary time (1–3 million years).
The relative importance of genetic and epigenetic inheritance remains a subject of considerable debate.
Darwin’s Finches and the Origin of Species
A reconstructed pattern of evolution (as drawn by Lack for Darwin’s finches) is known as a phylogeny. Two historical processes must have contributed to that phylogeny. Any evolutionary change in time – in anatomy, behaviour, development or molecular constitution – is known by the term phyletic evolution. If phyletic evolution had not occurred in the history of life, then all living things would look like their first ancestors. The second evolutionary process (or ‘mode’ of evolution) is ‘speciation’, the splitting in time of one species into two or more. If no speciation had occurred in the history of life, there would be, assuming a single common ancestor, only one species of living thing on Earth!
As we shall see later, there is still some dispute about the nature of speciation, particularly in animals. We have already seen that animal species are separated from one another by some sort of barrier, physical, behavioural or physiological, such that members of two different species do not normally interbreed with one another. This applies even in the case of two closely related (sibling) species occupying the same area, such as two species of Darwin’s finches living on the same small Galapagos Island. At some stage in their joint history a barrier to free interbreeding must have occurred. It has long been the claim of the evolutionary biologist Ernst Mayr that for that barrier to arise, splitting one species into two, there must have been an allopatric phase with the two, incipient species separated geographically. During the period that the two, incipient species are separated, differences in structure, behaviour, physiology, etc. evolve either in response to their different environments or by chance. If in the future the two species come to occupy the same habitat, the barrier to interbreeding will be enhanced by natural selection – Darwin’s great theory of the mechanism of evolution (see ‘The Galapagos Islands and Natural Selection’). Hybrids will either not be produced or will be at a competitive disadvantage to the pure-bred members of what are now separate species.
Darwin’s finches have long been regarded as ideal exemplars of allopatric speciation (isolated by a physical barrier). A little thought will demonstrate that there are two ways in which part of a single species can be separated from the rest:
1. If the range of a species is divided by some geographical event, such as the opening of a seaway or the origin of a mountain chain, free communication between members of the species on either side will cease. This is a vicariance event.
2. If on the other hand a sample of the population crosses a preexisting barrier, either voluntarily or by chance (wind-blown, or the floating mats of vegetation referred to in the section ‘The Galapagos Islands and Darwin’s Finches: A Case Study’, for example), then this is a dispersal event.
The islands were never connected to the South American mainland, from which they are far distant. They are also volcanic, so uninhabitable when first formed. But dispersal from the mainland to the islands is certainly possible if one considers that the major Galapagos fauna are birds and reptiles (mammals are rare):
Birds of course can fly. Many of the coastal nesting birds (e.g. albatross) are strong, long-distance flyers. Others could have been aided by prevailing winds.
Reptiles are particularly hardy animals and could theoretically survive for many months on floating vegetation mats.
Mammals are much more vulnerable to water loss and lack of food and therefore less likely to survive long sea journeys.
But the first dispersal event – South America to the Galapagos – does not explain the existence of 14 species on the islands. The finches are one of the best-known examples of adaptive radiation occurring due to allopatric speciation. Grant envisages:
1. The ancestral birds arriving on San Cristobal (Chatham).
2. Then after their adaptation to that island, a few birds ‘island-hopping’ to other islands to which they and their descendants became adapted.
3. The third and critical phase is when some island-hopping birds arrive on an island already inhabited by a finch population that has adapted to a particular way of life on that island. The new arrivals, if they survive, may have a different way of life and if breeding between the two stocks is inhibited, they will constitute sibling species descended from a common ancestor.
The whole process depends on many islands, at least slightly different in their environmental demands, and the rarity of island-hopping events over many generations. Without a sufficient time-frame, the bird stocks will not be sufficiently different for an interbreeding barrier to be formed – they will still be the same species. When the two stocks are established, reinforcement occurs. Reinforcement is due to any factor that causes the two species not to interbreed, and to occupy different ecological niches. The whole process can be summed up as divergence in allopatry and reinforcement in sympatry. Sympatry is where two or more species overlap as they diverge.
Thus, the theory of adaptive radiation by allopatric speciation implies that each species arose by differences in allopatry, presumably on a different island, and then reinforcement in (subsequent) sympatry. As a corroboration of this theory, evolutionary biologists point to the Cocos Island with its single finch species. There was no scope there for differentiation in allopatry after the ancestral birds had arrived.
It is a characteristic of good science that however firmly established a grand theory is, scientists themselves will question it. Newton’s mechanics were eventually superseded by Einstein’s relativity. Therefore, can we question the scenario of an adaptive radiation of Darwin’s finches on the Galapagos Islands? The answer is yes, of course, but in this case that questioning will lead to a deeper understanding of the historical process, rather than destruction of the theory.
(1) Until recently it was thought that no Galapagos island had ever been nearer to the South American mainland than those still existing. But in 1992, geologists discovered a series of seamounts (undersea extinct volcanic mountains) between the Galapagos and the mainland. Some of these underwater hills had cobbles indicating coastal erosion – they once protruded above the sea. So at least some island hopping between South America and the Galapagos might have occurred.
(2) For the extreme allopatric model, inter-island flight should be a rare event, but occasional vagrant species are seen on islands where they are not regarded as resident, and in recent years (1982–1983) the large ground finch has coloured the small island of Daphne Major.
(3) One case is known of the apparent beginnings of sympatric speciation, an island population dividing itself into two. The Island was Genovesa, small, remote and flat. The large cactus finch is resident on Genovesa and has been intensively studied by the Grants. They started their work there in 1978 after a severe drought in 1977. Male birds were heard to sing one of two distinct territorial songs, A or B. The nestling birds have one of two beak colours, yellow or pink. A males had 36% offspring with yellow beaks (and 64% with pink!), whereas B males had only 18% yellow. There was also a significant difference in bill length between adult A and B birds, correlated with different feeding habits. Type A birds fed on the flowers or hammered open the fruits of the prickly pear cactus, while B birds tore open the cactus pods searching for insects.
There seems little probability that either A or B birds had arrived from elsewhere. The nearest population of the large cactus finch to that on Genovese is the only other one known, and is on the distant island of Espanola about 200 km away. There is no evidence of any other population ever having existed between the two. In subsequent years, the correlation between song type and beak length disappeared and there was no evidence that females of type A parentage mated for preference with type A males (assortative mating). Nevertheless, the division of the population in 1978 suggests the initial stage of sympatric speciation.
(4) There have been many cases recorded of hybridisation between different species of Darwin’s finches, even those belonging to different genera. Over a period of 16 years, the Grants and their associates have conducted very detailed studies of the finches on the small crater island of Daphne Major. They discovered that the medium ground finches regularly hybridise at a low level with both the small ground finch and the small cactus finch, and that the hybrid offspring appear to be at no disadvantage, sometimes even flourishing more than pure-bred birds. This seems at odds with the idea that two sibling species in sympatry should evolve away from one another as the final stage of speciation. It also poses the question, too complex to deal with here, as to whether the six species of ground finches are in fact good species.
The Galapagos Islands and Natural Selection
It was Charles Darwin’s great achievement not just to suggest that evolution had occurred (and give cogent reasons for accepting this) but also to propose a mechanism that could produce evolutionary change. His theory of natural selection proposed such a mechanism, answering in part the question, how can evolutionary change, phyletic evolution, occur, and how is it that living organisms adapt to changes in their environment? Natural selection is a theory of adaptive change. And yet, as we have seen, it was not generally accepted that the differences in size and shape of the beaks in Darwin’s finches were adaptively significant until the publication of Lack’s book in 1947.
Darwin thought that natural selection, and thus adaptive change, was extremely slow and thus not open to observation. A study by Peter Boag and Peter Grant, published in 1981, showed that significant change can occur within a single generation. In 1977 the rains failed on the island of Daphne Major, resulting in very high mortality in the population of the medium ground finch, G. fortis, and no breeding. The birds that survived into 1978 were considerably larger than those that died (immediately showing the sex ratio, roughly equal before, of six males to one female). Seeds are the main food of G. fortis and in the drought year were in very short supply. This was particularly the case with the small seeds that were their staple. Only large birds with relatively deep beaks could open the seed cases and crush the seeds of the plant Tribulus. There was another plant with smaller seeds available, but this produced a sticky latex, gumming up the unfortunate bird.
Thus, selection for the ability to eat large tough seeds resulted in a change in mean beak depth, but this would not result in evolutionary change, unless the difference were heritable. To show that this was the case the beak depths of later offspring were plotted on a graph against the mean beak depth of each offspring’s parents demonstrating that beak depth is indeed heritable. Adaptive beak depth can result from natural selection.
A few years later the direction of selection was seen to reverse. In December 1982 the islands were hit by the effects of one of the most severe El Niño events of the twentieth century. The rains on Daphne Major continued to the end of summer and throughout the period G. fortis went into a breeding frenzy, each breeding pair, including some born in the same year, producing several broods. The population rose by some 400%. After the rains there came a population crash. But this time it was large birds (especially males) that were selected against. The reason is uncertain: there was in this case a surplus of small seeds relative to large, and it was suggested that, beak size or not, large birds simply needed more seeds, thus involving more searching to keep going. Also, it is said that the large beaks are less useful in the young before the beaks harden.
The sensational changes in one generation resulting from both events on Daphne Major impressed evolutionary biologists, but the Grant team themselves noted that what they had described was a case of stabilising selection. The birds on Daphne Major could react rapidly to climate change, but over the years, body and beak size fluctuated about a mean. There was no evidence of selection producing phyletic evolution – sustained directional evolutionary change. For natural selection to produce such change, there has to be new genetic information on which selection can act. We will discuss the origin of such information (known as mutation) in Chapter 3.
The Galapagos Islands therefore provide a powerful endorsement of Darwinian evolutionary theory along with an outstanding outdoor laboratory in which to study natural selection. The rate of evolution is rapid; in just under four million years there has emerged an amazing diversity of form arising from the animals and plants flying, floating, hitch-hiking or swimming there. Population sizes have become extraordinarily large in some cases, for instance in the unique marine iguana. Why should this be so?
One possible answer is in the relative lack of predators on the island. Snakes are small and ineffectual against all but the smallest reptiles. The Galapagos hawk is limited by nesting sites, and among the invertebrates the large (20 cm) centipede Scolopendra is a formidable adversary for other invertebrates but it rarely troubles the islands’ vertebrate population. Large jungle predators from the South American mainland (mainly mammals such as the jaguar) just could not make the journey. Also, visitors to the island frequently note how tame all the animals are. Such ‘island tameness’ is typical of those creatures not subject to extensive predation pressure.
It is suggested that a lack of predators coupled with smaller size and rapid reproduction (energy can now be diverted into reproductive success) has spurred the evolutionary trajectories of the Galapagos communities.
Perhaps disconcertingly, a point was made of featuring data that might conflict with orthodoxy. Does the discovery of seamounts to the east of the present archipelago cast doubt on the accidental dispersal of ancestors to the Galapagos? Does inter-island flight occur too frequently for the allopatric scenario to be valid? Does hybridisation between apparent species refute their states as good species? Could incipient sympatric speciation, as seen in the large cactus finch on Genovesa, go to completion? I suspect that the answer to all these questions is ‘no’, but they must be asked. Progress in science is made by the recognition of data that appear to contradict established theories, by debate between theories and by continuous, ruthless questioning.
Humans pollute – we can degrade the environment, but we also conserve. The last remaining giant tortoise on the island of Pinta, Chelonoidis abingdonii, was discovered in 1971 and relocated to the Charles Darwin Research Station on the island of Santa Cruz. Named ‘lonesome George’, this male Pinta giant tortoise was the very last of his kind. Attempts to locate other Pinta tortoises or mate him with females from closely related species all failed (although interestingly clutches of eggs were produced from such matings but none were viable). Sadly, Lonesome George died in 2012. The death of the ‘last of his kind’ was, of course, a blow but also a ‘wake-up call’ to conservationists globally. George’s death (at the relatively young age of 80 years or so) has reawakened conservation efforts both in the Galapagos and elsewhere resulting in a finding in 2015 that a closely related species, C. donfaustoi, had a 90% DNA match to that of George.